Primitive or Advanced? For a long time, trilobites were considered the most primitive of arthropods, since they were among the first Cambrian arthropods discovered, but this turns out to have been mistaken; an artifact of preservation. The first recorded Cambrian outcrops only preserved species with hard, shelly parts, such as trilobites, brachiopods, mollusks, and echinoderms. When the Burgess
Shale, Chengjiang, and other similar Cambrian Konservat-Lagerstätten (remarkably preserved fossil deposits) were discovered, large numbers of more delicate Cambrian arthropod species (that typically do not preserve at all) were revealed as contemporaries of trilobites, rather than their descendants (see diorama below). Trilobites are now considered relatively
advanced among the Paleozoic arthropods, and the search for the first arthropods and the ancestors
of trilobites is pushed back into the Precambrian.
Status of "Trilobitomorpha" In the 1959
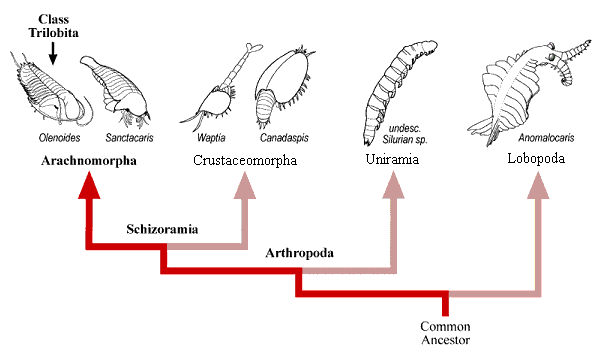
Trilobite Treatise, many of the Burgess Shale arthropods were included in the arthropod Subphylum Trilobitomorpha. Today that subphylum is no longer considered valid -- it served as a convenient unit for various paleozoic taxa with similarities in limb structure, but has fallen aside for more rigorous analyses. One prevalent recent classification recognizes the Trilobita as a class of arthropods sitting comfortably within the Superclass Arachnomorpha (an expanded concept based on the Chelicerata), alongside the Superclass Crustaceomorpha (based on an expansion of the Crustacea), together comprising the Subphylum Schizoramia (Arthropods bearing biramous limbs) contrasting with the Subphylum Atelocerata (insects, myriapods, and allies). This classification diagram is shown below.
TRILOBITA AMONG THE MAJOR CLADES OF PALEOZOIC ARTHROPODA
While there is certainly a great deal of diversity of form among the arthropods and near-arthropods (such as the large, predatory protarthropodan Anomalocaris), it seems reasonable that shared common
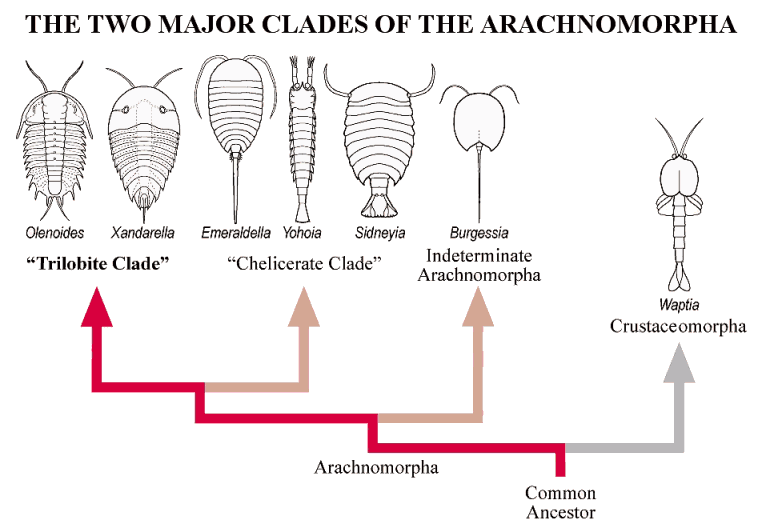
ancestry in the Pre-Cambrian was the basis for the radiations of the early Cambrian, such as seen at the Burgess Shale (Canada), Chengjiang (China) and Sirius Passet (Greenland). This shared ancestry and close relationship despite seemingly great divergence of form is seen in the very similar molecular biology of modern crustaceans and insects, and suggests to some workers that the Subphylum and Superclass designations for the Arthropoda may be superfluous (especially that dividing the Atelocerata from the Schizoramia). Arachnomorpha and Crustaceomorpha Among the diversity of Paleozoic arthropods, two large groups have emerged in recent phylogenetic analyses: a group of crustacean-like arthropods referred to as Crustaceomorpha, and a group referred to as Arachnomorpha. The diversity within the Arachnomorpha is dominated by Trilobita and Chelicerata, each forming major clades. Arachnomorpha (Størmer 1944) is equivalent to a grouping called Arachnata (Lauterbach 1983), which is defined as an inclusive grouping of non-crustacean arthropods: a clade stemming from the ancestor of Trilobita and Chelicerata. The Arachnomorph cladogram and its two major clades (sensu Cotton and Braddy 2004) is shown below.
“Chelicerate Clade” and “Trilobite Clade” Arachnomorph diversity allows attempts to elucidate the relationships between trilobites and other Paleozoic arachnomorphs (see, for example, Edgecombe & Ramskøld 1999, Cotton & Braddy 2004). It is relatively easy to exclude a large subset of arachnomorphs that fall into a “Chelicerate
Clade,” which can be generally characterized as arachnomorphs bearing a post-anal telson/tail, and a pair of limbs per segment (among other synapomorphies). The Burgess Shale arthropods Sidneyia and Yohoia are two good example members of this Chelicerate Clade (see above).
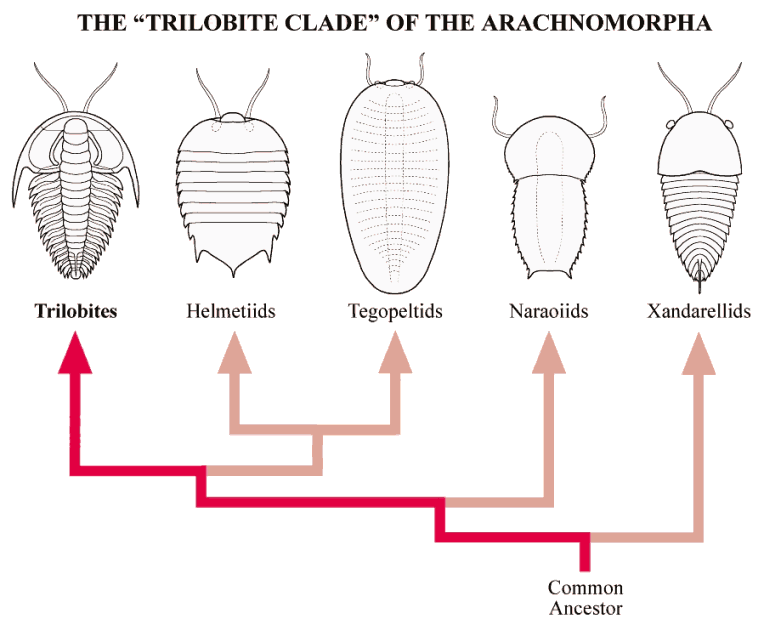
In contrast, the “Trilobite Clade” includes arachnomorphs lacking a telson, and with dorsal segments that may cover more than one pair of legs. The pygidium of trilobites is a good example of a fused single dorsal tergite covering several pairs of legs. Taken to extreme, the single-piece carapace of Tegopelte, and the two-piece carapace of Naraoia also represent this trend of decoupling of dorsal tergites with limb pairs (see below). We owe much of our recent understanding of the trilobite clade to the exceptionally well preserved assemblages of arthropods from Chengjiang, which greatly complements the assemblage from the Burgess Shale.
Features of the “Trilobite Clade” Most of the arachnomorph trilobite clade share features in common with trilobites, such as a hypostome, 3-4 pairs of post-antennular legs under the cephalon, similar limb structure, etc. Such similarities have led to proposals to include some trilobite-like arachnomorphs in the Class Trilobita (e.g., the Naraoiidae). However, some recent cladistic analyses of arachnomorph taxa have elevated Helmetiidae as a clade closely aligned with the trilobites (see figure above, and discussion below), which would create a paraphyletic situation if the naraoiids are also considered trilobites. It seems more reasonable to recognize the similarities of members of the “Trilobite Clade” of the Arachnomorpha, while also recognizing their distinctiveness.
Below, each of the members of the "Trilobite Clade" are discussed, and the constituent species are pictured. Several different sources were used to compile the reconstructions pictured here, and I am pleased to note that this site is the first publication (web or otherwise) to picture all of the pertinent genera and species of the "Trilobite Clade" in one place, and in a consistent style.
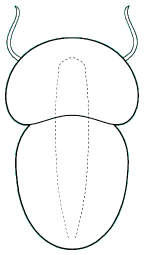
Helmetiidae
Helmetia expansa Burgess Shale
Kuamaia lata Chengjiang
Kuamaia muricata Chengjiang
Skioldia aldna Chengjiang
Rhombicalvaria
acantha Chengjiang
When the Burgess Shale animal Helmetia expansa was described (above, left), it was known from only a single, incompletely preserved specimen that displayed neither complete limbs nor antennae. The remarkable Chengjiang biota includes four species of helmetiids, clearly similar to Helmetia, and demonstrating antennae, trilobite-like limbs, stalked ventral eyes, and an anterior lobe. Most of the species show a distinct head shield, thorax, and tail shield, very similar to trilobites. The exception, Skioldia aldna, displays features intermediate between typical helmetiids and tegopeltids such as Saperion (described below). In several recent cladistical analyses, helmetiids emerge as the arachnomorph clade most similar to trilobites, displacing the Naraoiidae as the clade nearest to trilobites, and suggesting that the idea of naraoiids as a "ninth order" of trilobites should probably be set aside.
Kwanyinaspis maotianshanensis Chengjiang
In 2005 a Chengjiang soft-bodied arthropod was described with uncertain affinities. Kwanyinaspis
maotianshanensis Zhang & Shu 2005, was tentatively described as an aglaspid (a member of the "chelicerate clade"). However, Paterson et al 2010 suggested that Kwanyinaspis bears characters that suggest more affinities with the "trilobite-like clade." They place Kwanyinaspis as
basal
helmetiid, because of similarities in limb morphology and anteriorly reflexed thoracic segments. However, it also seems to bear characteristics of xandarellids (see below), especially in lack of a large tail shield, an axial posterior spine, lack of a discernable anterior lobe, and ventral eyes occupying dorsal bulges in the cephalon (as in Sinoburius).
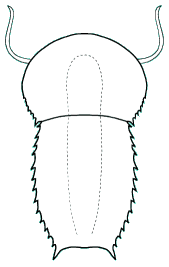
Tegopeltidae
Tegopelte gigas Burgess Shale
Saperion glumaceum Chengjiang
The large, enigmatic arthropod Tegopeltegigas from the Burgess Shale was initially described as a segmented "soft-bodied trilobite" akin to Naraoia. More recent reexamination of the specimen indicated the four purported tergite boundaries of Tegopelte were asymmetrical and probably compression fold artifacts. Saperion glumaceum, a similar large, ovoid arthropod with multiple somites covered by a single dorsal shield, was documented from the Chengjiang fossil beds, and was placed in the family Tegopeltidae in recognition of its similarity to Tegopelte. The ventral details of Tegopelte are poorly preserved, while specimens of Saperion clearly revealed stalked ventral eyes, and an anterior lobe similar to that found in helmetiids. My reconstruction of Tegopelte presumes that it also bears similar structures, although they are not clearly visible in the Burgess type specimen. Compelling similarities between Saperion and the helmetiid Skioldia (see above), particularly the splayed tergal boundaries that fail to reach the lateral margins, indicate the close relationship between helmetiids and tegopeltids. Indeed, the only diagnostic feature of tegopeltids is that the margin is entire, smooth, and unadorned by spines or serrations. It is clear that Helmetiidae and Tegopeltidae form a single monophyletic clade.
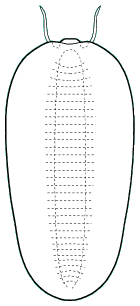
Naraoiidae
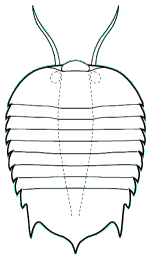
Naraoia compacta Burgess Shale
Naraoia spinifera Burgess Shale
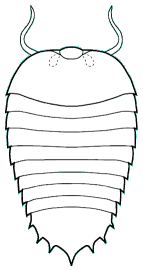
Naraoia spinosa Chengjiang
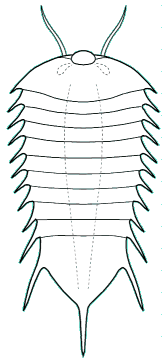
Naraoia
bertiensis Canada
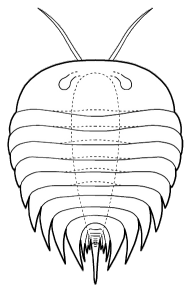
Misszhouia
longicaudata Chengjiang
The genera Naraoia and Misszhouia are very closely related and together comprise the family Naraoiidae. Misszhouia was originally classified as a member of the genus Naraoia, but was recently split out on the basis of its dissimilar antennal orientation (see above, right). When Naraoia compacta was described, its similarity to an early meraspid trilobite (with cephalon and pygidium, but no thoracic segments) was noted, and was part of an ontogenetic argument for including Naraoia as a "soft-bodied trilobite." A fuller discussion of the relationship of naraoiids to trilobites is presented on a separate
page of this website.
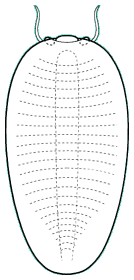
Liwiidae
Tariccoia arrusensis Puddinga Fm, Sardinia
Liwia convexa Cambrian, Poland
Soomaspis splendida Soom Shale, Africa
Buenaspis forteyi Sirius Passet
The family Liwiidae (sometimes subfamily Liwiinae of the family Naraoiidae) is comprised of naraoiid-like species that are distinguished from naraoiids in bearing thoracic tergites. Some, like Buenaspis (above, far right), are remarkably similar in general form to some advanced trilobites (e.g., Nileidae), but as in naraoiids, are primarily eyeless, and lack a well-defined axis (in addition to being uncalcified). The Liwiidae are geographically diverse, being recorded from a variety of sites outside of either the Burgess Shale or Chengjiang deposits, including one other Cambrian konservat-lagerstätte, Sirius Passet, North Greenland (where Buenaspis was found). Despite a diversity of form, the Liwiidae are considered most closely related to the Naraoiidae. However it remains to be seem whether the general similarities reflect true homologies.
Emucaridae
Kangacaris
zhangi Kangaroo
Island, Australia
Emucaris
fava Kangaroo Island, Australia
In 2010, a new family of Nektaspida, the Emucaridae, were described via two new genera of soft-bodied arachnomorphs from Australia's Kangaroo Island (Paterson et al 2010). This family is distinguished by bearing a small number of thoracic segments, with articulations poorly developed, and with no strong differentiation between either the cephalon in front nor the tail shield behind. This distinguishes the family from the Liwiidae, which typically show distinct cephalic, thoracic and tail shield morphology. The Emucaridae are similar to other arachnomorphs in several ways. Incomplete articulation of the tail shield is similar to that seen in tegopeltids, and the smooth transitions between cephalon, thorax and tail shield is similar to that seen in some xandarellids, such as Pygmaclypeatus (see below)
Xandarellidae
Xandarella
spectaculum Chengjiang
Cindarella
eucalla Chengjiang
Sinoburius
lunaris Chengjiang
Squamacula
clypeatus Chengjiang
Pygmaclypeatus
daziensis Chengjiang
The xandarellids include arachnomorphs with a well-defined head shield, stalked, ventral eyes (but sometimes furnished with dorsal portals, as in Xandarella and Sinoburius), and numerous dorsal tergites. The axis is either completely undeveloped (e.g., Squamacula, Pygmaclypeatus), or poorly defined (e.g., Sinoburius, Cindarella, Xandarella). Size of the tail shield varies considerably between species, and is quite trilobite-like in Sinoburius. Even where the tail shield is not well-developed, there is a decoupling of tergites and somites, so that posterior tergites cover increasing numbers of limbs (each limb pair corresponding to a somite). There are no xandarellids documented from the Burgess Shale. All the known species are from Chengjiang. They point to the value of this exceptional Cambrian konservat-lagerstätte, from which an entire clade of trilobite-like arachnomorphs was added to the fossil record.
Of course, no matter how similar the other trilobite clade arachnomorphs are to true trilobites in morphology, there are a number of features that trilobites share with no other arthropod group. These are discussed below:
What distinguishes Trilobites among Arthropods? Trilobites are the most diverse of the extinct arthropod groups, known from about 5000 genera (e.g., see Jell & Adrain 2003). The classification of trilobites within the Arthropoda has generated much controversy, much of which is still not completely resolved (see above). Whatever their higher position among arthropoda, there are a number of characters
that distinguish trilobites from within its Arachnomorph clade, the most significant noted below:
eye
ridges: These are consistently present in primitive trilobites, connecting the front of the palpebral lobe with the axial furrow (a feature lost in many post-Cambrian trilobites)
pygidium: The posterior tagma of greater than one segment is a conspicuous feature of all trilobites (but not restricted to Trilobita). Pygidia are typically very small in primitive forms (e.g., Olenellina)
Together with the organization of the body into three anterior-posterior divisions (cephalon, thorax, and pygidium), and the three longitudinal lobes (axial lobe and two flanking pleural lobes), the body features on this page serve to readily distinguish trilobites from all other known arthropod groups.
calcitic
compound eyes: While other compound eyes are found in Cambrian arthropods, only those of trilobites have corneal surfaces composed of prismatic calcite lenses (with the crystallographic axis normal to the lens surface).
circumocular sutures: In Cambrian holochroal trilobite eyes, a suture around the edge of the shared corneal surface assisted in molting of holaspid trilobites. In post-Cambrian trilobites this feature is secondarily lost, leaving the corneal surface attached to the librigena.
rostral plate: a ventral anterior plate separated from the rest of the cephalic doublure by sutures is very well developed in primitive trilobites (e.g., Redlichiida), narrower in other trilobite orders, and secondarily lost in some advanced forms (e.g., Asaphida and Phacopida)
hypostomal
wings: The trilobite hypostome may be homologous to the labrum in Crustacea, and other arachnomorphs bear hypostomes, but all trilobite hypostoma bear a pair of anterior wings which fit in pits in the anterior axial glabellar furrows (or homologous locations).
calcified cuticle: Trilobites bear a rather pure calcareous cuticle that ends ventrally at the inner edge of the doublure. Although a few other arthropod groups calcify, none do so the same way as trilobites. Crustacea, for example, are calci- fied ventrally and post-orally, so appendages are calcified.
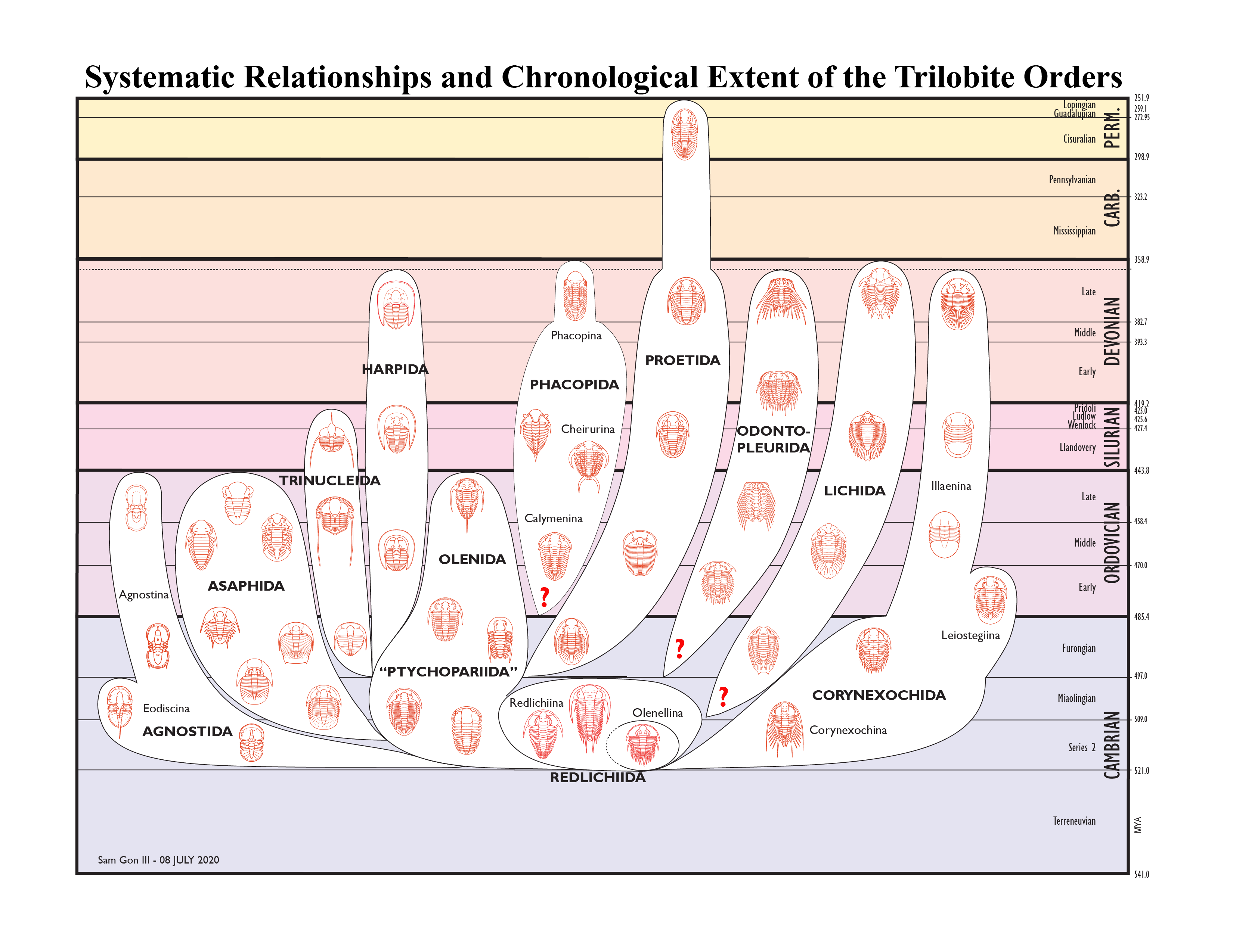
Relationships Between the Trilobite Orders The higher level classification of trilobites is an unsettled and dynamic subject, and since Fortey's classification outline in the 1997
Trilobite Treatise, we have seen his 2001 review of trilobite classification, the 2002 splitting of the Harpetida from the Ptychopariida, and recent acknowledgement of the orders Odontopleurida and Trinucleia suggests an arrangement that looks something like the figure below. A discussion of the evolutionary trends and relationships of the orders follows.
The Redlichiida (particularly the Suborder Olenellina) is considered primitive, appearing in Series/Epoch 2 of the Cambrian, and not persisting into the latest Cambrian (Furongian). Members of Ptychopariidaare
likewise among the first known trilobites. The Agnostida appear
thereafter, perhaps derived from Ptychopariida, and persist to the end of the Ordovician. The Redlichiina also give rise to the Corynexochida, in
the Cambrian. The Lichida and Odontopleurida may have arisen from early Corynexochida or Redlichiida, indicated with a "?". The Redlichiida, Corynexochida, and the Ptychopariida, as large primitive groups including the ancestors of other orders, must be paraphyletic. In 2002, another order was split out of the Ptychopariida; the Order Harpida (formerly suborder Harpina of the order Ptychopariida). In 2019, Order Trinucleida was distinguished from Asaphida. Order Olenida has been proposed, but a diagnostic description has not yet been offered in 2020.
The Ptychopariida, Harpida, Asaphida, and Proetida puportedly share (in at least the primitive forms) species with a natant hypostomal condition, leading Fortey to suggest the Subclass Libristoma for these orders combined. The recognition of Asaphida, Proetida, and Harpida as orders is a relatively recent thing; in the 1959 Treatise they were all included within the very large and paraphyletic Ptychopariida. The Ptychopariida and Harpida maintain the natant state until their extinction at the end of the Devonian, but both the Asaphida and the Proetida develop conterminant and impendent hypostomes in their advanced forms. The major extinction event at the end of the Ordovician greatly affected trilobites, ending the Olenida, Agnostida and Asaphida. The Trinucleida) are lost before the end of the Silurian and all other orders except Proetida are lost by the end of the Devonian (most in the major extinction event between the Frasnian and Famennian ages in the Late Devonian, but the Phacopidae hang on until nearly the Devonian-Carboniferous transition). The Proetida persist until the end of the Permian, the last of the orders of trilobites to go extinct. Only Order and Suborder epithets are provided above. There wasn't room for all of the superfamily figures and labels for the Proetida and Asaphida .
The origin of the Phacopida is uncertain. The three suborders (Phacopina, Calymenina, and Cheirurina) share a distinctive protaspis type; this similarity in development suggests phylogenetic closeness. The Calymenina is perhaps the most primitive of the Phacopida, and share some characters with the Ptychopariida/Olenida (including a few species with natant hypostomes), so although the hypostomal condition of the Phacopida is typically conterminant (and impendent in some advanced Phacopina), they may have had their origins with the natant Ptychopariida (which would make the Phacopida another addition to the Libristoma). Others point out the overwhelmingly conterminant hypostomal condition among Lichida, and similarities in the exoskeleton tuberculation of Phacopida and Lichida, so the ancestral sister group of the Phacopida remains unclear, although its clade in the chart above is placed nearest to Ptychopariida/Olenida.
In the first trilobite Treatise (1959), an Order Odontopleurida was recognized, including the large family Odontopleuridae. Thomas & Holloway (1988) proposed that the Lichidae and Odontopleuridae are related, but that their post-Cambrian evolutions have been distinct. When Fortey (1997) added Damesellidae to the order Lichida, it was indicated that they are more similar to Odontopleuridae than to Lichidae. In 2020 the two orders are recognized, and Lichidae+Lichakephalidae=Lichida, and Odontopleuridae+Damesellidae=Odontopleurida. However, the Cambrian family Lichakephalidae may be paraphyletic, with some of its genera similar to Lichidae and others more similar to Odontopleuridae. In particular, some workers reject the synonymy of Eoacidaspididae with Lichakephalidae, and note that at least some genera in Eoacidaspididae are close to Odontopleuridae.
notes on the use of this figure: This
diagram
was initially designed by Sam Gon III, based on information available in the literature, including the 1997 revision of the Treatise, and Fortey's 2001 synopsis of trilobite systematics. The intention is to share with others interested in trilobites my slowly growing understanding of the relationships between the higher taxonomic units. Any similarity to figures published elsewhere is unintentional, and I have not seen any handling of quite this sort in the literature. Please contact me before using this image. Any inaccuracies are entirely mine, and there may be future revisions. For example, since 1999, I adjusted the extinction of the Asaphida to the early Silurian, set the origin of the Phacopida to the Cambrian-Ordovician boundary, adjusted all of the Devonian extinctions to reflect the Frasnian-Famennian boundary, and added the small persistent tail on the Phacopida until its last family (Phacopidae) disappeared near the Devonian-Carboniferous boundary. When the Order Harpetida was recognized in late 2002, that order-level clade had to be added! In May of 2006, adjustments were made on the geological time scale to reflect the emerging treatment of the Cambrian, in which the appearence of trilobites is considered the bottom of Series/Epoch 2, and dashed lines indicating paraphyly in the primitive orders were added. In
2007,
working with Nigel Hughes, a modification of this figure was formally published (see citation for Hughes 2007, below). In 2008, the diagram was adjusted to split Odontopleurida out from Lichida. I intend to make continuing revisions to this figure as I receive feedback or learn more. Thank you for your understanding!
On what basis, then, are the orders defined? No
single character (e.g., facial sutures) dominates in higher level classification. Instead, such characters as facial sutures, glabellar shape and pattern of lobation, eyes, thoracic features and numbers of thoracic segments, pygidial shape, size and segmentation, and spinosity all play a role in helping define the orders. In addition, note that hypostomal conditions and shared ontogeny play an important role in defining the orders of trilobites. That is why for each of the order fact sheets, as many of the above details are provided as possible. As a synopsis, here are brief statements regarding each of the trilobite orders:
AGNOSTIDA - Among
the early trilobites, with a basic, clamshell-like appearance. Suborders Agnostina and Eodiscina. Representative species pictured here: Ptychagnostus
akanthodes (Agnostina)
REDLICHIIDA - Including
the most primitive trilobites from the lower Cambrian. Suborders Olenellina and Redlichiina. Representative species pictured here: Redlichia sp.(Redlichiina)
CORYNEXOCHIDA - An
often spiny group united by a shared hypostomal attachment. Suborders Corynexochina, Illaenina, and Leiostegiina. Representative species pictured here: Kootenia
sp. (Corynexochina)
.
ODONTOPLEURIDA - Very spiny trilobites, perhaps related o the Lichida. Suborder Odontopleurina; superfamilies Dameselloidea and Odontopleuroidea. Representative species pictured here: Selenopeltis
buchii.
LICHIDA - Some of the most ornately sculptured species fall into this group. Suborder Lichina; families Lichidae and Lichakephalidae. Representative species pictured here: Arctinurus
boltoni (Lichioidea)
PHACOPIDA- The well-known Phacops, with its beautiful compound eyes belongs here. Suborders Calymenina, Phacopina, and Cheirurina. Representative species pictured here: Phacops
sp.(Phacopina)
PROETIDA - Includes some of the last trilobite species before the Permian Extinction. Suborder Proetina, with three Superfamilies. Representative species pictured here: Proetus
granulosus (Proetoidea)
TRINUCLEIDA - Mostly bearing a wide fenestratd fringe and long, narrow genal spines, a distinct group split from Asaphida by Bignon et al 2019.
Representative species pictured here: Cryptolithus sp.
ASAPHIDA - All share a ventral median suture, and most a similar development. Suborder Asaphina, with several Superfamilies comprising ~20% of all trilobites. Representative species pictured here: Homotelus
sp. (Asaphoidea)
PTYCHOPARIIDA - Bearing the "generic trilobite" body plan, but many weird variations! Suborders Ptychopariina and Olenina (Harpina has been elevated to order Harpetida; see below) Representative species pictured here: Modocia
sp. (Ptychopariina)
HARPIDA - Bearing the distinctive, broad, often intricately pitted, cephalic fringe. In 2002, split out of the Ptychopariida and elevated from suborder to full order. Representative species pictured here: Eoharpes sp.
NEKTASPIDA - The so called "soft-shelled trilobites" such as Naraoia have been classified as an order of trilobites by some. Click on the image or link to learn more about them, and to see how they are handled in the 1997 Treatise.
If you would like to see a listing of the families of each of the orders in phylogenetic arrangement according to the 1997 Treatise and other sources, please click here. Literature Cited:
Bignon, A., B. G. Waisfeld, N. E. Vaccari, & B.D.E. Chatterton. 2019. Reassessment of the Order Trinucleida (Trilobita). J. Syst. Paleontol. 18(13): 1061-77.
Chen, J. & G. Zhou. 1997. Biology of the Chengjiang Fauna. in The
Cambrian Explosion and the Fossil Record. Bulletin of the National Museum of Natural Science 10:11-106.
Cotton, T.J., and S.J. Braddy. 2004. The phylogeny of arachnomorph arthropods and the origins of the Chelicerata. Transactions of the Royal Society of Edinburgh: Earth Sciences, 94: 169–193,
Ebach, M.C. & K.J. McNamara. 2002. A systematic revision of the family Harpetidae (Trilobita). Records of the Western Australian Museum 21: 235-67.
Edgecombe, G. & L. Ramskøld. 1999. Relationships of Cambrian Arachnata and the systematic position of Trilobita. J. Paleontology. 73(2):263-87.
Fortey, R.A. 1997. Classification. In Kaesler, R. L., ed. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. xxiv + 530 pp., 309 figs. The Geological Society of America, Inc. & The University of Kansas. Boulder, Colorado & Lawrence, Kansas.
Fortey, R.A. 2001. Trilobite systematics: The last 75 years. J. Paleontology. 75(6) 1141-51.
Lauterbach, K.-E. 1983. Synapomorphien swischen Trilobiten- und Cheliceraten-sweig der Arachnata. Zoologischer Anzweiger 210:213-38.
Paterson, J.R., G.D. Edgecombe, Diego C. Garcia-Bellido, J.B. Jago & J.G. Gehling, 2010. Nektaspid arthropods from the Lower Cambrian Emu Bay Shale lagerstatte, South Australia, with a reassessment of lamellipedian relationships. Palaeontology 53(2):377-402
Størmer, L. 1944. On the relationships and phylogeny of fossil and recent Arachnomorpha. A comparative study on Arachnida, Xiphosurida, Eurypterida, Trilobita, and other fossil Arthropoda. Skrifter Utgitt av Det Norske Videnskaps-Academi I Oslo. I. Matematisk-Naturvidenskapelig Klasse 5:1-158.
Wills, M.A., D.E.G. Briggs, R.A. Fortey, M. Wilkinson & P.H.A. Sneath. 1998. An arthropod phylogeny based on fossil and Recent taxa. In:
G.D. Edgecomb, ed. Arthropod Fossils and Phylogeny. Columbia University Press, N.Y.
Zhang X.L. & D.G. Shu. 2005. A new arthropod from the Chengjiang Lagerstatte, Early Cambrian, southern China. Alcheringa
29:185-194.





 Image
above ©2020 by S. M. Gon III, created using Adobe illustrator
Image
above ©2020 by S. M. Gon III, created using Adobe illustrator