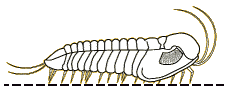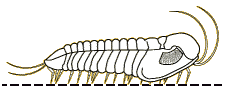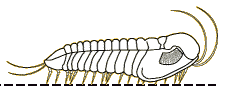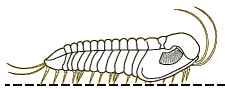
Profallotaspis jakutensis
SIBERIA
| Profallotaspis jakutensis SIBERIA | The International Commission on Stratigraphy has proposed that the start of Series 2 of the early Cambrian is marked by the first appearance of trilobites in the fossil record. Depending on the dating system, this would be about 521 million years ago, on what are currently the continents of Africa ( |
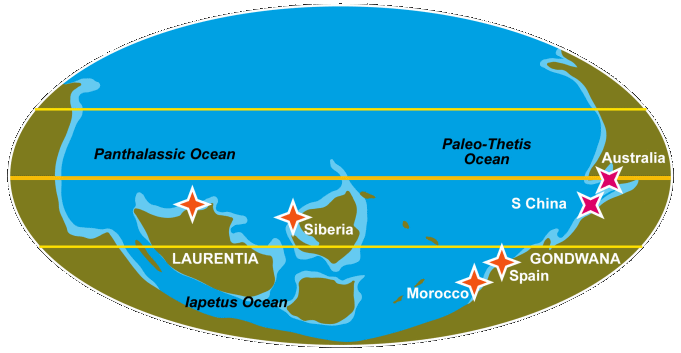
Early Cambrian paleocontinental positions and the first trilobite localities.
Regional stratigraphy of the first trilobites
In Siberia, the first trilobites mark the base of the Atdabanian Stage. Profallotaspis is succeeded by two species of Repinaella, as well as Bigotina. As noted above, this set is most similar to the western Laurentian assemblage. The next zone is characterized by Delgadella (previously Pagetiellus) with Archaeaspis hupei at the base. These early trilobites appear long after archaeocyaths and brachiopods in the same area (Repina, 1981). In Laurentia, the first trilobites appear below what was once considered the base of the Montezuman Stage, marked by the "Fallotaspis Zone." This Fritzaspis Zone is named for the first appearance of the trilobite Fritzaspis, a form lacking genal spines, which is rapidly followed by Profallotaspis?, Amplifallotaspis and Repinaella? which together comprise the Fritzaspis assemblage. In Morocco, the first trilobites appear simultaneously with archaeocyaths, with Hupetina appearing first, Eofallotaspis appearing just afterward (Geyer, 1996; Sdzuy 1978). In Spain, bigotinids appear at a similarly early position (Pillola, 1993; Liñan et al., 2005). The first trilobites in south China and in Australia, both species of Abadiella (redlichiids), appear to be younger than the early trilobites of Siberia, Morocco, and western Laurentia (Paterson and Brock, 2007). Additional radiometric dates -- there being none from the Cambrian of western Laurentia -- and geochemical studies should resolve these correlations. For the remaining global regions, the first trilobites include Schmidtiellus in Baltica, and Callavia in western Avalonia. But both are distinctly younger than the old trilobites of Siberia, Morocco, Spain, and Laurentia.
The first trilobite genera - DRAFT -
Very shortly after the appearance of the first trilobites, there are several other genera that appear in each of the regions. Here is a listing of some of these early genera, in rough order of appearance, oldest at bottom. The three tiers of this table roughly correspond to the Lower, Middle, and Upper Atdabanian of Siberia. The boldfaced genera are the nominate genera for descibed biostratigraphic zones (e.g., the Fritzaspis Zone of Laurentia, the Eofallotaspis Zone of Morocco, etc.).
| LAURENTIA | SIBERIA | SPAIN | MOROCCO | AUSTRALIA | CHINA |
| Grandinasus Esmeraldina Palmettaspis Nevadia "Nevadella" Cambroinyoella Montezumaspis | Bonnia Uktaspis Triangulaspis Delgadella Judomia | Granolenus Delgadella Eoredlichia | Antatlasia | Abadiella | Hupediscus Sinodiscus Tsunyidiscus Abadiella |
Bradyfallotaspis? "Cirquella" Parafallotaspis Fallotaspis Archaeaspis | Paranevadella Bigotinella "Nevadella" Delgadella Archaeaspis | Lemdadella Bigotina | Daguinaspis Choubertella Lemdadella Fallotaspis Eofallotaspis | no trilobites | no trilobites |
| Repinaella? Amplifallotaspis Profallotaspis? Fritzaspis | Repinaella Profallotaspis | Serrania bigotinid | Eofallotaspis Hupetina | no trilobites | no trilobites |
The first trilobite orders
Note that within the small set of first trilobites above, four orders are already represented: Redlichiida (represented by fallotaspidoids, redlichioids, and olenelloids), Ptychopariida (represented by ellipsocephaloids), Agnostida (represented by eodiscoids), and somewhat later, Corynexochida (represented by corynexochoids). What this really suggests is that early in trilobite evolution, the distinguishing features between the orders were not clearcut, and trilobites very close to the basal ancestors of those early lineages were developing characteristics that might be applied to several orders. At this early stage of evolution, the orders must be paraphyletic, if not polyphyletic. The similarities can be seen by this comparison of the cephala of a few representative early trilobites:
| Fritzaspis LAURENTIA | Profallotaspis SIBERIA | Serrania SPAIN | Hupetina MOROCCO | Eofallotaspis MOROCCO |
|
|
|  |
|
|
| Archaeaspididae Fallotaspidoidea | Archaeaspididae Fallotaspidoidea | Bigotinidae Ellipsocephaloidea | Bigotinidae Ellipsocephaloidea | Fallotaspididae Fallotaspidoidea |
All have parallel-sided or tapering glabellae with simple, even lobation, and well-developed, wide-set eye ridges that converge on and typically merge into the anterior lobe of the glabella. There is often a wide preglabellar area (indicating a natant hypostome). There may or may not be genal spines present, but when present, they can be well-developed. Another notable tendency is for there to be a rearward pointing angle in the posterolateral margins of the cephalon, sometimes bearing a metagenal spine. Some have suggested that the ridges and furrows running from the ends of the ocular ridges to the metagenal angle or spine in fallotaspidoid trilobites are homologous with the posterior facial sutures of suture-bearing trilobites.
Facial sutures in early trilobites
From the very start of the trilobite record, facial sutures are present in bigotinids. In Spain and Morocco (Western Gondwanaland) Lemdadella is another early trilobite bearing sutures. Jell (2003) noted that sutures seem to appear independently in another major early lineage: the redlichiids of Australia and China, but it is not at all clear from what ancestor they arose. It is quite interesting that the first trilobite found in
Early trilobite evolution
Jell (2003) offered several considerations on the early evolution of trilobites, presenting evidence that the primitive condition for hypostome attachment was natant, not conterminant as proposed by Fortey (1990). In that case, that primitive natant condition is maintained in the Redlichiida and the Ptychopariida, while the conterminant condition becomes the primary state for derived orders such as Lichida, Corynexochida, Asaphida, and Phacopida. Jell also offered an origin of the Agnostida via the Bigotinidae, demonstrating similarities between both the glabellar and pygidial characters of meraspid bigotinids and holaspids of early eodiscines such as Tsunyidiscus, and proposing an origin of the eodiscines via progenesis, retaining bigotinid meraspid morphology while attaining reproductive maturity. He also offered early evolutionary pathways for the origins of the primitive Asaphida and Corynexochida.
Very shortly after trilobites appeared in the lower Cambrian, they rapidly diversified into the major Cambrian orders: Redlichiida, Ptychopariida, Agnostida and Corynexochida. These orders in turn proliferated, diversified, and gave rise to the rich variety of forms that trilobite workers have described in ten orders, well over 175 families, over 5,000 genera, and over 17,000 species, persisting for a quarter of a billion years before their extinction at the end of the Paleozoic Era.
Fortey, R. A. 1990. Ontogeny, hypostome attachment, and trilobite classification. J. of Paleontology. 33:529-76.
Geyer, G. 1996. The Moroccan fallotaspidid trilobites revisited. Beringeria, 18, 89-199.
Hollingsworth,
J.S., 2008. The first trilobites in Laurentia and elsewhere. In: I.Rábano, R. Gozalo and D. García-Bellido (Eds.), Advances in trilobite research. Cuadernos del Museo Geominero, nº 9. Instituto Geológico y
Jell, P. 2003. Phylogeny of Early Cambrian trilobites. In: Lane, P.D., Siveter, D.J. and Fortey, R.A. (eds.), Trilobites and their relatives, contributions from the Third International Conference, Oxford, 2001. Special Papers in Palaeontology, 70. The Palaeontological Association,
Liñán, E., R. Gozalo, M. E. Dies Álvarez, J.A. Gámez Vintaned & S. Zamora. 2008. Nuevos trilobites del Ovetiense inferior (Cámbrico Inferior bajo) de Sierra Morena (España) [New Lower Ovetian Trilobites (Low Lower Cambrian) From Sierra Morena (Spain).] Ameghiniana 45(1)
Pillola, G.L. 1993. The Lower Cambrian trilobite Bigotina and allied genera. Palaeontology, 36, 855-881.
Sdzuy, K. 1978. The Precambrian-Cambrian boundary beds in Morocco (preliminary report). Geological Magazine; March 1978; v. 115; no. 2; p. 83-94.