| . . Olenelloidea
Olenellus
Fallotaspidoidea
Fallotaspis
 |
Suborder Olenellina Cephalon: lacking facial sutures; glabella typically with rather deep lateral furrows; in some species the front glabellar lobe is an almost circular boss; natant or conterminant hypostome, very wide rostral plate extending between genal angles, with perrostral suture (no connective sutures).
Thorax: with numerous segments, non-fulcrate; axis often spine-bearing.
Pygidium: narrow, with few segments.
Other: fallotaspidoids are among the earliest trilobites in the Cambrian; no calcified protaspis known among olenelloids; body flattened; cuticle thin.
Superfamilies: Olenelloidea (2 families), Fallotaspidoidea (5 families) Superfamily Olenelloidea
Cephalon: glabellar outline concave, anterior glabellar lobe (LA) usually enlarged, glabella narrowest at L2 or S1, L3 usually modified distally with posterolateral part bending backward and encroaching on L2; ocular lobe connects only to posterolateral part of LA. Compare to Fallotaspidoidea (below).
Thorax: as in Olenellina.
Pygidium: as in Olenellina.
Other: appearing after the Fallotaspidoidea in Laurentia
Families: Olenellidae, Holmiidae
Genera: Olenellidae: Angustolenellus, Arcuolenellus, Biceratops, Bolbolenellus, Bristolia, Fremontella, Fritzolenellus, Gabriellus, Laudonia, Lochmanolenellus, Mesolenellus, Mesonacis (=Fremontia), Mummaspis, Nephrolenellus, Olenelloides, Olenellus (/Barrandia), Paedeumias, Peachella, Teresellus, Wanneria.
Holmiidae: Andalusiana, Baltobergstroemia, Callavia (=Cephalacanthus/Callavalonia; =Cobboldus), Cambropallas, Elliptocephala (/Georgiellus, /Ebenezeria), Holmia (=Esmeraldina), Holmiella, Iyouella, Kjerulfia, Palmettaspis, Postfallotaspis, Schmidtiellus (/Schmidtia). Superfamily
Fallotaspidoidea
Notes: Includes some of the oldest known trilobites (Fritzaspis, Eofallotaspis, Profallotaspis).
Cephalon: glabellar outline cyclindrical or slightly conical, L3 simple, similar to L2; occular lobe usually connects at anterolateral part of LA. Compare to Olenelloidea (above).
Thorax: as in Olenellina.
Pygidium: as in Olenellina.
Families: Fallotaspididae, Archaeaspididae, Judomiidae, Neltneriidae, Nevadiidae.
Genera: Archaeaspididae: Archaeaspis, Bradyfallotaspis, Fallotaspidella, Fritzaspis, Geraldinella, Profallotaspis, Repinaella, Selindella.
Fallotaspididae: Choubertella, Daguinaspis (=Eodaguinaspis; =Epidaguinaspis), Eofallotaspis, Fallotaspis, Lenallina, Parafallotaspis, Pelmanaspis, Wolynaspis.
Judomiidae: Judomia, Judomiella, Paranevadella, Sinskia.
Neltneriidae: Bondonella, Neltneria.
Nevadiidae: Buenellus, Cambroinyoella, Cirquella, Limniphacos, Nevadella, Nevadia, Plesionevadia, Pseudojudomia, Sdzuyomia.
|
Emuelloidea
Balcoracania
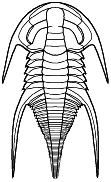
Redlichioidea
Redlichia
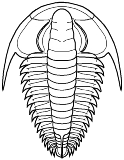
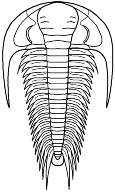
Paradoxidoidea
Paradoxides
 | Suborder Redlichiina Cephalon: with opisthoparian facial sutures; early forms tend to have tapering, conical glabella with furrows extending far backwards; later forms with glabella expanding forwards to inflated frontal lobe; hypostome conterminant (e.g., Redlichia) or natant (e.g., Dolerolenus), rostral plate narrower than in Olenellina, bound by rostral and connective sutures.
Thorax: fulcrate or non-fulcrate, typically with many segments (60+ in an Emmuelid); axis infrequently spine-bearing.
Pygidium: typically small, but can be larger and with many segments.
Superfamilies: Emuelloidea, Redlichioidea, Paradoxidoidea. Superfamily Emuelloidea
Cephalon: cranidium subquadrate, glabella cylindrical, slightly contracted at S3, 3 pairs of glabellar furrows, preglabellar field short or absent, eye ridge wide, long, directed slightly posterolaterally, palpebral lobe cresentic, posterior area of fixigena with fulcrum, librigena with long genal spine; hypostome conterminant, attached to narrow rostral plate.
Thorax: divided into prothorax of 3 or 6 segments (with last prothoracic segment macropleural) and long opisthothorax of up to 95+ segments (Balcoracania holds the record for greatest number of thoracic segments in a trilobite species).
Pygidium:.a minute, segmented disc.
Families: Emuellidae, Megapharanaspidae
Genera: Emuella, Balcoracania, Megapharanaspis. Superfamily Redlichioidea
Cephalon: as in Redlichiina, but not with characters of Emuelloidea or Paradoxidoidea.
Thorax: as in Redlichiina, but not with characters of Emuelloidea or Paradoxidoidea.
Pygidium: as in Redlichiina, but not with characters of Emuelloidea or Paradoxidoidea.
Other: largest and most diverse superfamily of Redlichiina.
Families: Redlichiidae, Dolerolenidae, Yinitidae, Mayiellidae, Gigantopygidae, Saukiandidae, Metadoxididae, Abadiellidae, Kuechowiidae, Menneraspididae, Redlichinidae, Chengkouaspidae.
Genera: Abadiellidae: Abadiella (=Parabadiella; =Danagouia), Guangyuanaspis, Guangyuania, Lunolenus, Malongocephalus, Shaanxia, Sibiriaspis.
Chengkouaspidae: Aragotus, Bathynotus (=Pagura), Bathynotellus, Belliceps, Chengkouaspis, Elegestina, Inella, Pseudoresserops, Terechtaspis (=Nellina).
Dolerolenidae: Dolerolenus (/Olenopsis; =Malungia), Giordanella, Granolenus, Paramalungia.
Gigantopygidae: Bornemannaspis, Gigantopygus, Parayiliangella, Pseudoyiliangella, Yilliangella (=Palaeoaspis), Yilliangellina, Zhangshania.
Kueichowiidae: Kueichowia, Shatania.
Mayiellidae: Mayiella, Qiaodiella, Qiaotingaspis
Menneraspididae: Menneraspis.
Metadoxididae: Churkinia, Conomicmacca, Enantiaspis, Fuminaspis, Hongshiyanaspis, Metadoxides (=Anadoxides), Minusinella, Onaraspis, Pratungusella.
Redlichiidae: Breviredlichia, Chaoaspis, Chengjiangaspis, Conoredlichia, Elganellus, Eoredlichia (=Archaeops; = Saukiandops; =Galloredlichia; = Pararedlichia), Hesa, Iglesiella, Irgitkhemia, Jingyangia, Kepingaspis, Kuanyangia, Latiredlichia, Lemdadella, Leptoredlichia (=Paraleptoredlichia), Maopingaspis, Metaredlichia, Mianxianella, Nebidella, Neoredlichia, Ningqiangaspis, Olgaspis, Pachyredlichia, Parawutingaspis, Parazhenbaspis, Pseudoredlichia, Pseudowutingaspis, Pteroredlichia (=Spinoredlichia), Redlichia (/Hoeferia; =Mesodema; =Dongshania), Redlichops, Sapushania, Sarassina, Sardaspis, Sardoredlichia, Syndianella, Tolbinella, Ushbaspis (=Metaredlichioides), Wengangaspis, Wutingaspis, Xela, Xenoredlichia, Yorkella, Zhanglouia.
Redlichinidae: Asthenaspis, Kolbaspis, Parasajanaspis, Redlichina, Sajanaspis, Sekwiaspis, Tungusella.
Saukiandidae: Australaspis, Clariondia, Despujolsia, Dolerolichia, Eops, Ezhimia, Longianda, Pareops, Perrector (=Rawops), Planocephalus, Pseudosaukianda, Realaspis, Resserops, Richterops (=Marsaisia), Saukianda.
Yinitidae: Drepanopyge, Drepanuroides (=Xishuiella), Hongjunshaoia, Longduia, Meitanella, Paokannia, Parapaokannia, Parayinites, Pseudopaokannia, Qingkouia (=Paradrepanuroides), Yinites, Yunnanaspidella, Yunnanaspis.
Family uncertain:
Akbashichia, Fandianaspis, Iolgia, Micangshania, Xingzishania. Superfamily Paradoxidoidea
Cephalon: semicircular cephalon with long librigenal spine, glabella widens anteriorly to rounded or bluntly pointed LA (parallel in early forms), L1 - L4 equal or subequal, S1 generally transglabellar, but shallow medially, S2 long, may be transglabellar, S3 and S4 short (S4 strongly oblique in Centropleuridae, making large angle with S3), preglabellar field generally absent in adult, opisthoparian, anterior facial sutures divergent, transverse, to retrodivergent.
Thorax: with 14 - 21 segments, short to long pleural spines directed progressively more strongly backward from front to rear, sometimes tips bluntly truncated (Centropleuridae), 1st and 2nd segments may be macropleural.
Pygidium: generally small in Paradoxididae, but tending to medium sized in Centropleuridae.
Other: often large trilobites.
Families: Paradoxididae, Centropleuridae, Xystriduridae.
Genera: Paradoxididae: Acadoparadoxides (/Entomolithus/Entomostracites; =Eoparadoxides), Anabaraceps, Anabaraspis, Bajanaspis, Baltoparadoxides, Phanoptes (=Eccaparadoxides; =Macrocerca), Hydrocephalus (=Phlysacium; =Rejkocephalus), Paradoxides (=Vinicella), Plutonides (/Plutonia), Primoriella, Schagonaria, Schoriina.
Centropleuridae: Anopolenus, Beishanella, Centropleura, Clarella, Luhops.
Xystriduridae: Galahetes, Inosacotes, Polydinotes, Xystridura (/Milesia). |
| ADDITIONAL CLASSIFICATION NOTES FOR REDLICHIIDA:
As one of the most primitive order of trilobites (along with Ptychopariida), the two suborders, Redlichiina and Olenellina, each pose some taxonomic problems. The Redlichiina may be considered the most basal group of non-olenellid trilobites, alongside the ellipsocephaloid Ptychopariida. Much of the historical taxonomy of Redlichiida has been developed stratigraphically. Both ptychopariid and corynexochid trilobites have sister taxa that could currently be classified as Redlichiida, making the order paraphyletic. For example, the Ellipsocephaloidea, once considered part of the Redlichiida, is now considered a primitive "bridge" between the Redlichiida and Ptychopariida. As sister taxa for other orders are defined in the Redlichiina, an evolutionarily meaningful classification may emerge.
Geyer (1996) in his review of Moroccan fallotaspidids questioned the importance of lack of facial sutures uniting the Fallotaspidoidea with Olenelloidea in the suborder Olenellina, pointing out that Fallotaspis and Lemdadella are difficult to distinguish if facial sutures are disregarded, and that although olenelloids follow fallotaspidoids stratigraphically, there is no clear transition between any fallotaspidoids and olenelloids, whereas holmiid olenelloids resemble redlichioids such as Gigantopygus. Geyer suggested that at least some olenelloids may have arrived at their lack of facial sutures secondarily from redlichioids.
Jell (2003) examines the earliest lineages of trilobites and has some controversial proposals. He notes that the earliest trilobites were probably natant (vs Fortey's assertion that an attached hypostome is the primitive state). This means that some lineages went from natant to attached, and others may have remained natant (leading to the Librostoma). Another assertion is that facial sutures may have arisen from fallotaspidoids more than once, and that one lineage (from Profallotaspis to Archaeaspis to Judomiidae and Holmiidae to Olenellidae) retained the primitively sutureless condition to give rise to the Olenelloidea. If Jell's lineages are valid, then both suborder Redlichiina and order Ptychopariida are polyphyletic! Indeed, recent research on the oldest trilobites in the fossil record indicate that in portions of Gondwanaland (now parts of Spain and Morocco), the first trilobites are ellipsocephaloids. This further points to the very close relationship between the Ellipsocephaloidea and Redlichiida. With a primitive lack of facial sutures, and a stratigraphically early range, the Olenellida have been recognized as distinct sister taxon to "typical" trilobites, and arguments have been made to exclude them from the Trilobita. Fortey (1990) argued effectively for maintaining the Olenellina as trilobites, but the relationship between the Olenellina and Redlichiina and other suture-bearing trilobites is by no means clear. They seem united by shared primitive characters (e.g., micropygy, numerous thoracic segments with spinose tips, long, cresentic eyes, etc.). Without a more clarified relationship, the Order Redlichiida as now defined (including both Redlichiina and Olenellina, and excluding Ellisocephaloidea) may not be justified. Jell (2003) suggests that Eofallotaspis gives rise to Lemdadella, and
thence to Eoredlichia and the Redlichiidae.
The ontogeny of olenelloids shows an interesting detachment of the hypostome from the rostral plate in mid-ontogeny, with a subsequent redocking of the hypostome by maturity. This may be relevant to the evolution of the natant orders (Ptychopariida, etc.), but is also consistent with Jell's (2003) suggestion that the natant state is primitive.
The family Bathynotidae was included in the order Redlichiida in the 1959 Treatise, but is listed as "order uncertain" in the 1997 Treatise. Its dorsal features are easily placed in the Redlichioidea, but its ventral features (particularly rostral plate lacking/greatly reduced) defy the typical pattern. Jell & Adrain (2003) nonetheless place Bathynotus in the redlichioid family Chengkouaspidae, with which it shares many dorsal characters. More recently, in 2013 Elicki and Geyer placed new species of the redlichioid genus Myopsolenites in the family Bathynotidae. They argued that Myopsolenites and the entire Onaraspis clade belongs in the Bathynotidae. This would suggest that the family would not merit a subordinal placement, but instead sit among the primitive redlichioid families discussed above, closest, perhaps, to the Metadoxididae. The placement of fallotaspidoid genera in the families Archaeaspididae and Fallotaspididae in Jell & Adrain is not universally accepted, with arguments for placement of Profallotaspis and Repinaella in Archaeaspididae by Geyer (1996) and Hollingsworth (2008). Fritzaspis Hollingsworth, is placed in Archaeaspididae. Paterson & Edgecombe removed Holyoakia from the Emuellidae (and in fact, removed it from Order Redlichiida, arguing that it is a member of Dorypygidae, Order Corynexochida). Paterson & Jago 2006 described Megapharanaspis, a new genus of emuelloid.
Elicki, O. & G. Geyer. 2013. The Cambrian trilobites of Jordan--taxonomy, systematic and stratigraphic significance. Acta Geol. Polonica 63(1):1-56.
Fortey, R. A. Trilobite systematics: the last 75 years. J. of Paleontology. 75(6):1141-51.
Fortey, R. A. 1990. Ontogeny, hypostome attachment, and trilobite classification. J. of Paleontology. 33:529-76.
Geyer, G. 1996. The Moroccan fallotaspidid trilobites revisited. Beringeria 18:89-199.
Hollingsworth,
J.S., 2008. The first trilobites in Laurentia and elsewhere. In: I.Rábano, R. Gozalo and D. García-Bellido (Eds.), Advances in trilobite research. Cuadernos del Museo Geominero, nº 9. Instituto Geológico y Minero de España, Madrid.
Jell, P.A. 2003. Phylogeny of Early Cambrian trilobites. Special Papers in Palaeontology 70:45-57.
Jell, P.A. & J.M. Adrain. 2003 Available generic names for trilobites. Memoirs of the Queensland Museum 48(2):331-553
Paterson, J.R. & G.D., Edgecombe. 2006. The Early Cambrian trilobite family Emuellidae Pocock, 1970: Systematic position and
revision of Australian species. Journal of Paleontology 80(3): 496-513.
Paterson, J.R. & J.B. Jago. 2006. New trilobites from the Lower Cambrian Emu Bay Shale Lagerstätte at Big Gully, Kangaroo Island,
South Australia. Memoirs of the Association of Australasian Palaeontologists 32:43-57. |