. Introduction to the Bathynotidae The small, trilobite family Bathynotidae is known from late lower Cambrian and the earliest Middle Cambrian sites in North America and Eurasia. This is a time in early Paleozoic history when the orders of trilobites were restricted to the Redlichiida, Ptychopariida, Agnostida, and Corynexochida. The 1959 Treatise placed the family in a suborder of the Redlichiida (Bathynotina). The 1997 Treatise treats the Bathynotidae as a family of "uncertain order." What is it about the Bathynotidae that leads to this uncertain status? Given what we know of the characters of the orders, why is it that the Bathynotidae confound ordinal placement? This page explores the morphological characters of the Bathynotidae, dorsal and ventral, in search of an explanation.
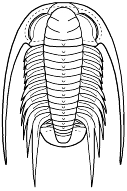
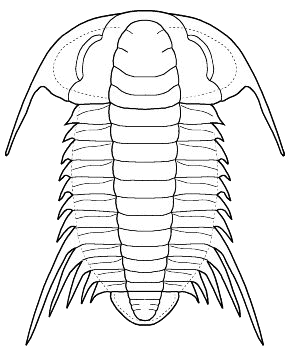
Bathynotus holopygus
Bathynotus holopygus, Hall 1860 E. North America &Arctic Eurasia
Redlichiine dorsal characters? Based on dorsal morphology alone (see figure, left), the Bathynotidae share many characteristics with members of the Order Redlichiida, Suborder Redlichiina, Superfamily Redlichioidea. The glabella is simple and tapering forward, eyes long and cresentic, long genal spines, thoracic axiswide, with 13 spinose thoracic segments, 11th segment macrospinose. . This set of characters is not unheard of in the Redlichioidea. A wide axis is seen in the Saukiandidae. Several redlichioid families bear 12 to 14 thoracic segments (Menneraspididae bear exactly 13). Macrospinosity is seen in the Metadoxididae (e.g., Onaraspis) and Saukiandidae (indeed Despuljosia is notably similar in spine pattern to Bathynotus). The dorsal facial sutures of Bathynotus are opisthoparian, with the anterior sutures converging in arcs, and merging smoothly with an anterior marginal suture. This is also seen among a few Redlichioids (e.g., Enantiaspis and Onaraspis (Metadoxididae)). There is little or no preglabellar area in Bathynotus, and a few redlichioids share this (e.g., Hamatolenus, Saukiandia, and Perrector). The eyes are large and cresentic, as in many Redlichiida (and Corynexochida). The spinous nature of Bathynotus is also shared by early trilobites in both the Redlichiida and Corynexochida. Many of these features are plesiomorphic, defining the "primitive" set of characters applying broadly to the paraphyletic Redlichiida. The row of axial nodes on Bathynotus can be found in Redlichiida, Corynexochida, and even Ptychopariida.
. The general similarity of form between Bathynotus and a selected set of redlichioid species is rather easily seen below. Based on the dorsal morphology alone, it would be reasonable to place Bathynotus in one of the advanced redlichioid families, such as Saukiandidae or Metadoxididae. Jell & Adrain (2003. Available generic names for trilobites. Memoirs of the Queensland Museum 48(2):331-551) place Bathynotus in the Redlichioid family Chengkouaspidae.
Bathynotus holopygus Bathynotidae
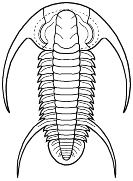
Despujolsia rochi Saukiandidae
Perrector (Richterops) falloti Saukiandidae
Onaraspis somniurna Metadoxididae
Specific similarities to Bathynotidae are noted in the various captions of the redlichioid taxa to the right. Compare to Bathynotus above.
Cresentic eyes, similar pattern of thoracic spines, including a macrospinose 11th segment followed by subspinose segments.
Glabella wide, tapering, simple; macrospinose segment followed by subspinose segments, axial nodes, pygidium simple, multisegmented.
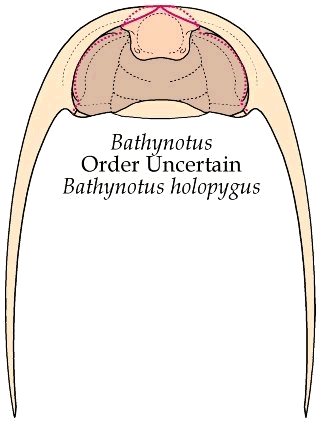
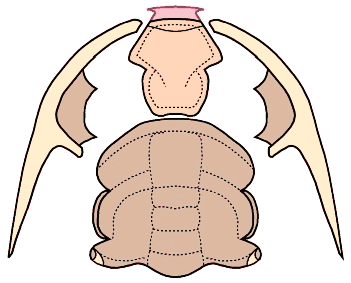
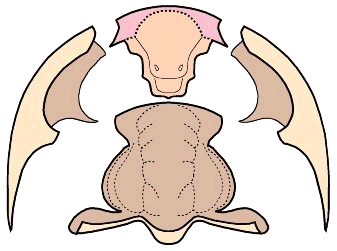
"Unique" ventral cephalic morphology While the dorsal morphology of Bathynotus is not hard to ally with that of Redlichioidea, the ventral morphology of Bathynotidae, as shown by Bathynotus holopygus (see images below), is a different matter entirely. It is apparently unique among trilobites. The 1997 Treatise goes so far as to state: "No other species is known with such a ventral structure, or with a triangular rostral plate in which the apex of the triangle is directed forward." The hypostome is conterminant, that is, it meets the anterior cephalic doublure and its placement is congruent with the anterior edge of the overlying glabella. This much is typical of primitive trilobites (Redlichiida and Corynexochida), but unlike other Redlichiida (which typically bear wide rostral plates), there is no obvious rostral plate in Bathynotus, and the sutures that separate the hypostome from the anterior doublure diverge at a roughly 100° angle from the anterior midline, giving the hypostome a roughly pentagonal shape (see below, left). In his review of the hypostomes of Cambrian trilobites, Whittington (1988) suggested that the triangular area anterior of the median body in the hypostome of Bathynotus may represent a remnant of the rostral plate, fused with the hypostome, which would make the two sutures with the anterior doublure connective sutures (e.g., separating the doublure from the rostral plate, rather than separating doublure from hypostome). There is certainly no clear indication of a suture between the purported rostral remnant and the hypostome proper, and even so, in nearly all other trilobites with small rostral remnants, the typical pattern is for the sutures to diverge forwards, rather than converge toward an apex. Thus, while the dorsal features of Bathynotus sit comfortably in the Redlichioidea, its ventral features seem difficult to reconcile. .
In the image below, the dorsal facial sutures are shown as dotted red lines, and the unusual anterior sutures of the hypostome are shown in solid red. Most lower Cambrian trilobites bear wide rostral plates that dominate the anterior doublure.
The explode view of the hypostome, librigenae and cranidium of Bathynotus show clearly the pentagonal shape of the hypostome. The apex of this pentagon may represent a remnant, fused rostral plate, but this is by no means clear.
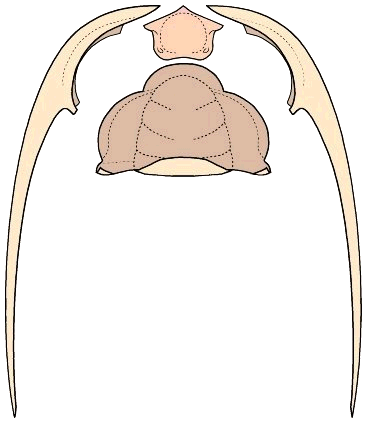
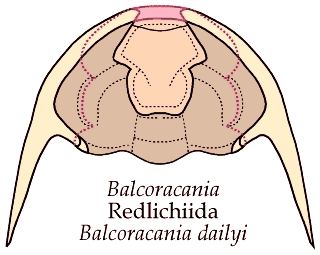
What trilobites come closest to Bathynotus in ventral structure? While no trilobites exactly match the lost or greatly reduced rostral plate of Bathynotus, among trilobites with conterminant hypostomes bearing small rostral plates there are two examples in which the rostral sutures converge anteriorly. The first example is in the Redlichiida, in the Emuelloidea. The ventral structure of the Australian emuellid Balcoracania
dailyi is shown below. The rostral plate (pink) is not fused with the hypostome (orange), but bears a functional suture. The two connective sutures initially converge anteriorly, then diverge to meet the dorsal sutures along the anterior margin. .
In the image below, the dorsal facial sutures are shown as dotted red lines, and in solid red lines are the connective sutures on either side of the rostral plate (pink). The hypostome is attached to the rostral plate by a functional transverse suture. The rostral plate of Balcoracania is narrower than the hypostome.
The explode view of the hypostome and rostral plate, librigenae and cranidium of Balcoracania show some similarities to that of Bathynotus. It is not too difficult to imagine the rostral plate shrinking until it is essentially gone. In that case, the connective sutures would have to meet at the anterior midline as in Bathynotus.
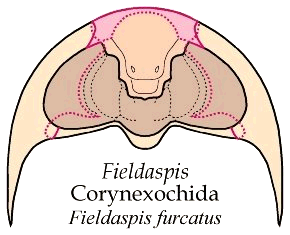
The other example is in the trilobite Fieldaspis furcatus, A trilobite in the order Corynexochida, Family Zacanthoididae. The details of the structures of this trilobite are shown below. As is apparent, the rostral plate (pink) is fused with the hypostome (orange), so that the median body marks the border between the hypostome and the rostral plate (dotted red line). The two rostral sutures converge anteriorly, but meet the dorsal sutures along the anterior margin, which bears a marginal suture. If the right and left rostral sutures were to migrate medially, they would converge on the hypostome and create an apex as shown in Bathynotus.
In the image below, the dorsal facial sutures are shown as dotted red lines, and in solid red lines are the connective sutures on either side of the rostral plate (pink). The hypostome is fused with the rostral plate without any sign of a functional transverse suture. The rostral plate of Fieldaspis is rather narrow, being just slightly wider than the hypostome.
The explode view of the fused hypostome-rostral plate, librigenae and cranidium of Fieldaspis show some similarities to that of Bathynotus. It is not too difficult to imagine the rostral portion of the fused hypostome-rostral plate shrinking until it is essentially gone. In that case, the connective sutures would have to meet at the anterior midline.
Considering the full complement of characters shown by Bathynotus, the dorsal features do not seem to justify placement in the Corynexochida, while its ventral features show characters that might be considered compatible with a few Corynexochida or Redlichiida, as shown above. It is particularly interesting that another redlichiine, Balcoracania (albeit in the Superfamily Emuelloidea, rather than Redlichiioidea), bears a ventral suture arrangement that is arguably similar to that of Bathynotidae. If any of the ventral structures of the advanced redlichioids (say, Saukiandidae) were shown to be similar to Bathynotus, that would argue for placement of Bathynotidae in the Redlichiida. Unfortunately, our knowledge of the hypostomes and ventral sutures of Redlichiida are limited, so it will be difficult to investigate for related taxa. For example, the hypostomes and ventral sutures for redlichioids that seem dorsally most similar to Bathynotus (e.g., Despuljolsia, Perrector, Onaraspis) are not known. If their ventral sutures should prove similar to Bathynotus, perhaps placement of Bathynotidae in the Redlichiida is reasonable. Similarly, the ventral and hypostomal structures of members of the Chengkouaspidae should also be investigated.
In 2013 Elicki and Geyer described Cambrian trilobites of the Burj Formation in Jordan, and placed new species of the redlichioid genus Myopsolenites in the family Bathynotidae. Their discussion of the affinities of Myopsolenites with Bathynotus included very similar pygidial form, a macropleural thoracic segment followed by two ultimate segments, and a hypostome fused to the rostral plate (as presumed in Bathynotus) with slightly medially directed rostral sutures. They argued that Myopsolenites and the entire Onaraspis clade belongs in the Bathynotidae. This would suggest that the family would not merit a subordinal placement, but instead sit among the primitive redlichioid families discussed above, closest, perhaps, to the Metadoxididae.
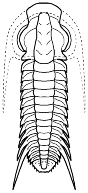
Irvingella sp. Elviniidae Ptychopariida
Ptychopariida connections? If Bathynotus resembles some advanced Redlichioids, it also has been compared to some of the Ptychopariida, such as Irvingella
(left). Comparing the spine patterns, facial sutures, glabellar structure, and pygidial structure of Irvingella (and other Elviniidae/Komaspididae) to Bathynotus is very easy to do, and it has even been argued that the Bathynotidae are the ancestors of the Komaspididae. Shergold, J.H. & Whittington, H.B. (2000) in The Cambrian trilobite Bathynotus (?Redlichioidea) in the Northern Territory, Australia. Alcheringa 24:1-10, state that "Kobayashi (1935) allied Bathynotus with Upper Cambrian forms such as Komaspis and Irvingella in his new family Komaspididae (in the Ptychopariida). Later, Hupe (1953), while acknowledging that Bathynotus may have been ancestral to komaspidids, proposed the family Bathynotidae and placed it with the Redlichioidea.
Trevor Cotton (in a trilobite discussion thread, March 2003) suggested recently that "Bathynotus is likely to belong to a clade of trilobites including some higher redlichioids, corynexochoids, ptychoparioids (and the other libristomates) and paradoxidoids. Bathynotus shares characters with some redlichioids, corynexochoids and paradoxidids." He noted that "a close relationship between Ptychopariida and Corynexochiida is a long-standing hypothesis in trilobite phylogeny (based e.g. on protaspis morphology, among other things). The similarity in dorsal morphology of adult Ellipsocephaloidea and corynexochids supports this. The corynexochid-ptychopariid clade originated from "advanced redlichioids" - and I think there has been some discussion of which redlichioids may be close to these more modern trilobite groups." Given the position of Bathynotus among the diversification of the Corynexochida, the Ptychopariida, and the advanced Redlichioidea, Cotton suggests finally that "the advanced redlichioids (probably including Bathynotus) could not form a clade excluding these groups," as they are descendants.