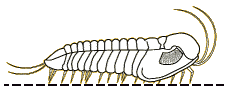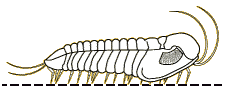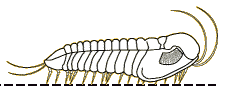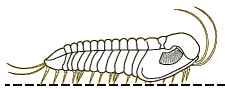
A
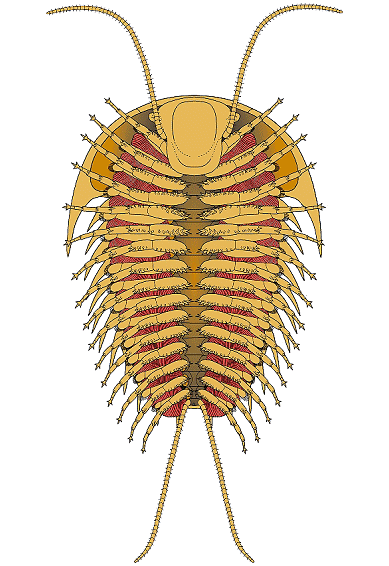
2003 paper by Nigel Hughes reviewed all of the trilobite species
with
descriptions of ventral features, presenting a summary table
including
the data below. It indicated that the typical trilobite bore a
pair of
antennae, then 3 cephalic limbs, followed by trunk (thorax +
pygidium)
limbs of variable number, depending on the number of thoracic
and
pygidial segments. The typical limb consisted of 6 or 7
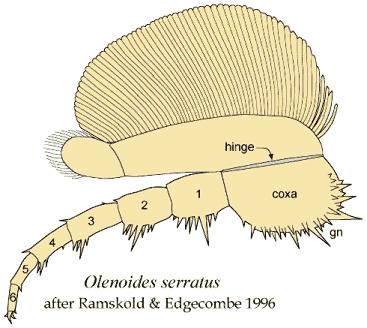
podomeres. Olenoides serratus
remains the only trilobite with antenniform posterior cerci
preserved. In 2017 Zeng et al provided an update on species
bearing ventral preservation, which is adapted here:
| Species |
Age |
Locality |
Cephalic
limbs |
Thoracic
limbs |
Pygidial
limbs |
Podomeres |
Source |
| Eoredlichia
internedius |
E. Cambrian |
Chengjiang |
3? |
n/a |
n/a |
?7 |
Shu et al 1995; Ramskold &
Edgecombe 1996 |
| Yunnanocephalus
yunnanensis |
E. Cambrian |
Chengjiang |
4? |
n/a |
n/a |
n/a |
Shu et al 1995 |
| Hongshiyanaspis yiliangensis |
E. Cambrian |
Kunming |
4? |
|
|
7 |
Zeng et al 2017 |
| Olenellus getzi |
E. Cambrian |
Lancaster, PA |
n/a |
n/a |
n/a |
n/a |
Dunbar 1925 |
| Olenoides serratus |
M. Cambrian |
Burgess Shale |
3 |
7 |
<6 |
6 |
Whittington 1975, '80 |
| Kootenia burgessensis |
M. Cambrian |
Burgess Shale |
n/a |
n/a |
n/a |
n/a |
Walcott 1918; Raymond 1920 |
| Elrathina cordillerae |
M. Cambrian |
Burgess Shale |
n/a |
n/a |
n/a |
n/a |
Walcott 1912, '18; Raymond
1920 |
| Elrathia
permulta |
M. Cambrian |
Burgess Shale |
n/a |
n/a |
n/a |
n/a |
Walcott 1918; Raymond 1920 |
| Agnostus pisiformis |
L. Cambrian |
Orsten, Sweden |
3 |
2 |
3 |
7 |
Muller & Walossek 1987 |
| Placoparia cambriensis |
M. Ordovician |
S. Wales |
3/3.5 |
n/a |
n/a |
n/a |
Whittington 1993; Edgecombe
& Ramskold 1999 |
| Isotelus
latus |
L. Ordovician |
. |
n/a |
?8 |
n/a |
n/a |
Raymond 1920 |
| Isotelus maximus |
L. Ordovician |
. |
n/a |
?8 |
?16 |
n/a |
Raymond 1920 |
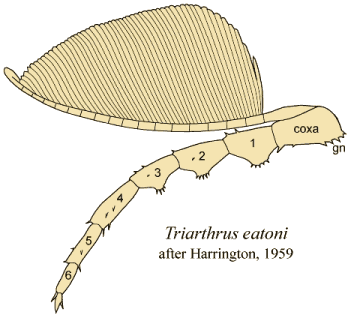
| Triarthrus eatoni |
L. Ordovician |
Beecher Trilobite Bed |
3/3.5 |
14 |
>10 |
6 |
Raymond 1920; Cisne 1973;
Whittington & Almond 1987; Edgecombe &
Ramskold 1999 |
| Cryptolithus
tesselatus |
M.-L. Ordovician |
. |
n/a |
?5 |
>10 |
?7 |
Raymond 1920; Stormer 1939 |
| Primaspis trentonensis |
M.-L. Ordovician |
. |
n/a |
?10 |
n/a |
n/a |
Raymond 1920; Ross 1979 |
| Primaspis
sp. |
L. Ordovician |
. |
n/a |
>8 |
n/a |
n/a |
Ross 1979 |
| Ceraurus
pleurexanthemus |
L. Ordovician |
Walcott-Rust |
4 |
?11 |
3 |
n/a |
Walcott 1918, '21; Raymond 1920;
Stormer 1939, '51 |
| Flexicalymene senaria |
L. Ordovician |
Walcott-Rust |
n/a |
?13 |
>2 |
n/a |
Walcott 1918, '21; Raymond 1920 |
| Chotecops ferdinandi |
E. Devonian |
Hunsruck |
3 |
11 |
>12 |
6 |
Sturmer & Bergstrom 1973;
Bruton & Haas 1999 |
| Asteropyge sp. |
E. Devonian |
Hunsruck |
3 |
?11 |
>?4 |
n/a |
Sturmer & Bergstrom 1973 |
| Rhenops cf.
anserinus |
E. Devonian |
Hunsruck |
3/3.5 |
11 |
>6 |
7 |
Bergstrom & Brassel 1984;
Bartels et al 1998; Edgecombe & Ramskold 1999 |
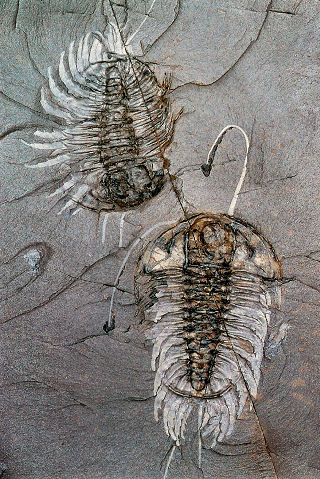
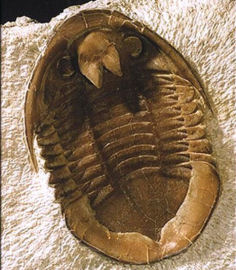
Here is an example specimen of Rhenops
cf. anserinus from Hunsruck, Germany, that has been
prepared to exposed both dorsal and ventral surfaces.
Limbs and antennae are preserved in pyrite.
Image courtesy of Andreas Ruckert:
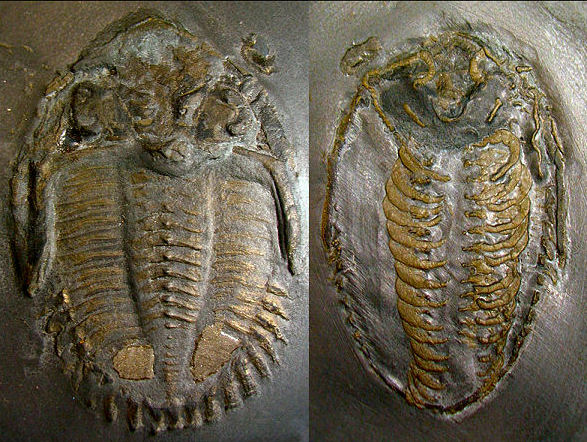
At the 2008 Trilobite
Conference in Spain, this image of the New York trilobite
Triarthrus eatoni won
wide acclaim as the best preserved cephalic ventral features of
a
trilobite yet found. The arrangement of the cephalic limbs
converging
on the hypostome indicate how the gnathobases act as mouthparts
processing and manipulating food in the vicinity of the mouth,
which
underlies the hypostome. Image courtesy of the collection of Dr.
Ed
Staver:

Bartels, C., D.E.G. Briggs, and G. Brassel. 1998. The
Fossils of the Hunsruck slate: Marine life in the Devonian.
Cambridge Paleontological Series Number 3. Cambridge University
Press, Cambridge.
Bergstrom, J. & G. Brassel 1984. Legs in the trilobite Rhenops
from the lower Devonian Hunsruck Shale. Lethaia
17:67-72.
Bruton, D.L. & W. Haas 1999. The anatomy and functional
morphology of Phacops
(Trilobita) from the Hunsruck Slate (Devonian). Palaeontographica
Abteilung A 253:1-75.
Cisne, J.L. 1973. Life history of an Ordovician trilobite Triarthrus
eatoni. Ecology
54:135-42.
Dunbar, C.O. 1925. Antennae in Olenellus
getzi, n. sp. Amer.
J. of Science. Series 5. 9:303-8.
Edgecombe, G.D. & L. Ramskold 1999. Relationships of
Cambrian Arachnata and the systematic position of Trilobita.
J. Paleontol. 73:263-87.
Hughes, N.C. 2003. Trilobite tagmosis and body patterning from
morphological and developmental perspectives. Integr.
Comp. Biol. 43:185
Muller, K.J. & D. Wallossek 1987. Morphology, ontogeny, and
life habit of Agnostus
pisiformis from the Upper Cambrian of Sweden. Fossils
and Strata 19:1-124.
Ramskold L. & G.D. Edgecombe 1996. Trilobite appendage
structure -- Eoredlichia
reconsidered. Alcheringa 20:269-76.
Raymond P.E.1920. The appendages, anatomy, and relationships of
trilobites. Memoirs of
the Connecticut Academy of Sciences 7:1-169.
Ross, R.J. Jr. 1979. Additional trilobites from the Ordovician
of Kentucky. United States
Geological Survey Professional Paper 1066-D:1-13.
Shu, D., G. Geyer, L. Chen, and X. Zhang. 1995. Redlichiacean
trilobites with preserved soft-parts from the lower Cambrian
Chengjiang
fauna (South China). Berlingeria,
Special Issue 2:203-41.
Stormer, L. 1939. Studies on trilobite morphology, Part I. The
thoracic appendages and their phylogenetic significanceNorsk.
Geol. Tidssk. 19:143-274.
Stormer, L. 1951. Studies on trilobite morphology, Part III.
The ventral cephalic sutures, with remarks on the zoological
position
of the trilobites. Norsk.
Geol. Tidssk. 29:108-58.
Stürmer, W. & Bergström, J. 1973: New
discoveries on trilobites by X-rays. Paläontologische
Zeitschrift
47, 104–141.
Walcott, C.D.1912. Cambrian geology and paleontology II. No. 6.
Middle
Cambrian Branchiopoda, Malacostraca, Trilobita, and
Merostomata. Smithsonian
Miscellaneous Collections 57:145-228.
Walcott, C.D. 1918. Cambrian geology and paleontology IV. No.
4. Appendages of trilobites. Smithsonian
Miscellaneous Collections 67:115-216.
Walcott, C.D.1921. Cambrian geology and paleontology IV. Notes
on structure of Neolenus.
Smithsonian Miscellaneous Collections 67:365-456.
Whittington, H.B. 1975. Trilobites with appendages from the
Middle Cambrian Burgess Shale, British Columbia. Fossils
and Strata 4:97-136.
Whittington, H.B. 1980. Exoskeleton, moult stage, appendage
morphology, and habits of the Middle Cambrian trilobite Olenoides
serratus. palaeontology 23:171-204.
Whittington, H.B. 1993. Anatomy of the Ordovician trilobite
Placoparia. Phil. Trans. R.
Soc. London. series B 339:109-18.
Whittington, H.B. & J.E. Almond 1987. Appendages and habits
of the Upper Ordovician trilobite Triarthrus
eatoni. Phil. Trans. R. Soc. London. series B 317:1-46.
Zeng, H., F. Zhao, Z. Yin, and M. Zhu. 2017. Appendages of an
Early Cambrian metadoxidid trilobite from Yunnan, SW China
support mandibulate affinities of trilobites and artiopods. Geol.
Mag. 154(6):1306-28.
|
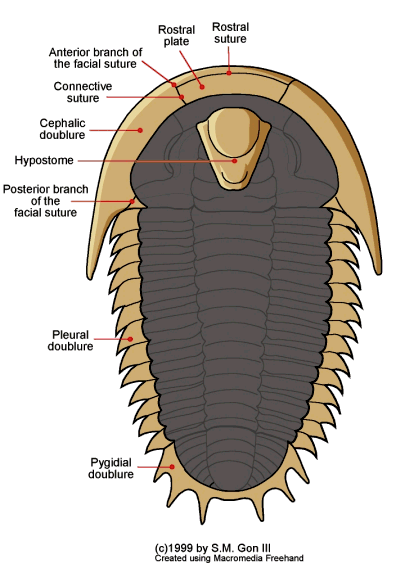
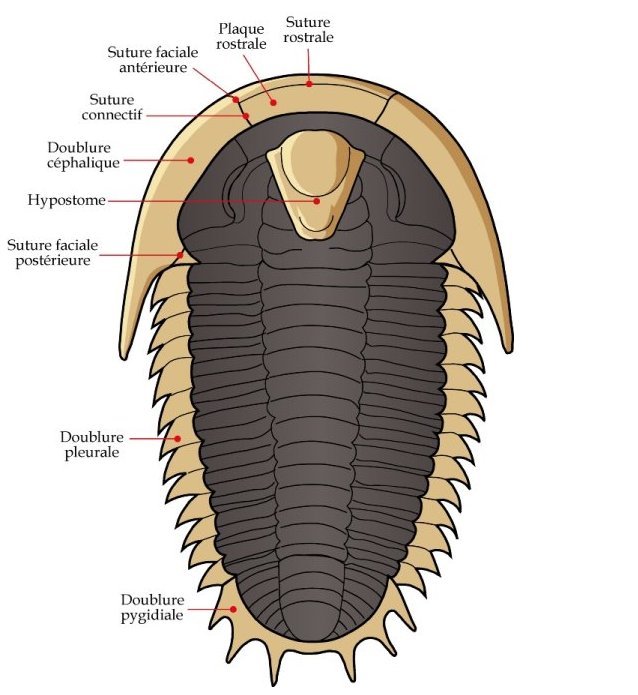
 Unlike the thicker dorsal shell of a trilobite, many of the
ventral (underside) features, including limbs and antennae,
usually are not preserved. The
ventral portions that are
typically preserved include the doublure
(a ventral extension of the dorsal exoskeleton), a special part
of the doublure, typically separated by sutures at the anterior
of the cephalon, called the rostral
plate, and a hard mouthpart called the hypostome,
that typically underlies the glabella.
Unlike the thicker dorsal shell of a trilobite, many of the
ventral (underside) features, including limbs and antennae,
usually are not preserved. The
ventral portions that are
typically preserved include the doublure
(a ventral extension of the dorsal exoskeleton), a special part
of the doublure, typically separated by sutures at the anterior
of the cephalon, called the rostral
plate, and a hard mouthpart called the hypostome,
that typically underlies the glabella.