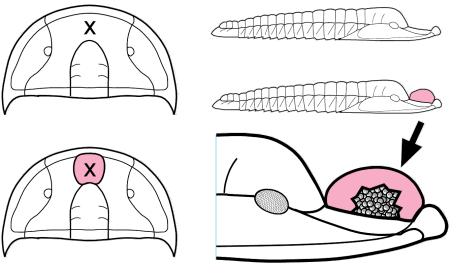
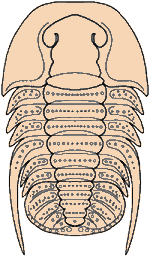
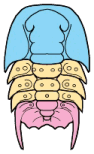
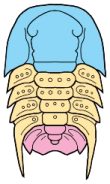
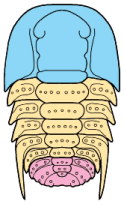
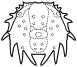
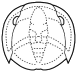
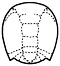Trilobites are thought to have reproduced sexually, as do nearly all arthropods today. Eggs were presumably laid, but fossilized eggs that may be of Cambrian eodiscid trilobites have been documented only once (Zhang and Pratt 1994). It has recently been suggested that some trilobites may have held eggs and/or developing young within the cephalon (as horseshoe crabs do today), and anterior median swellings of the cephalon (of the preglabellar field) in some specimens are interpreted as brood pouches (e.g., Fortey & Hughes 1998) because they appear only in holaspids (adults) and represent a dimorphism in which the swelling is the only morphological difference. Ostracods and some other crustaceans show similar brood pouch swellings, although not at the anterior of the body. Because specimens with brood pouches appear only in natant trilobites, it is possible that the eggs or protaspids were released ventrally, anterior of the hypostome.
Brood Pouches in Trilobites see Fortey, R.A. & N.C. Hughes. 1998. Brood pouches in trilobites. J. Paleontol. 72(4):638-49.
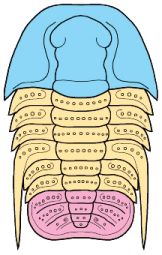
Left: dorsal view of cephalon of adult holaspid male; preglabellar field is marked "X" and female brood pouch swelling of preglabellar field is colored (lower left). Right: side view of male specimen (top), brooding female (middle) and close-up cut-away showing developing eggs within pouch.
Above: photo of specimen showing brood pouch
The embryology of trilobites is unknown, but after hatching and development of a hard exoskeleton (which presumably occurred at or soon after hatching) they molted their exoskeletons as they grew, as all arthropods today, leaving a record of their developmental stages. Thus, the ontogeny (patterns of growth and development) of trilobites is known only as far as calcite exoskeletons of immature stages have sometimes been preserved, recording the progression of forms that trilobite larvae and juveniles take from hatching to maturity. If any of the earliest stages lack a calcite exoskeleton, these would also escape our knowledge. To illustrate the ontogeny of trilobites, consider Dimeropyge (Order Proetida, Superfamily Bathyuroidea), shown below (via Chatterton, in the Treatise on Invertebrate Paleontology, 1997):
Protaspid period
Meraspid period
Holaspid period
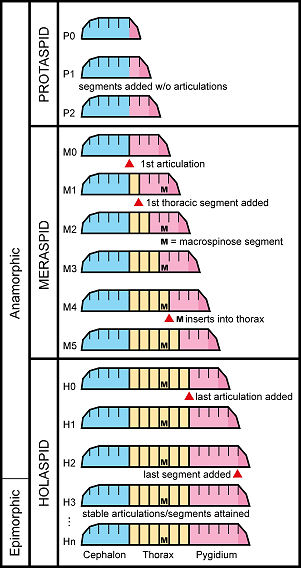
Three developmental periods are recognized: a protaspid period, meraspid period, and a holaspid period. In the protaspid period, the larva (called a protaspis) is composed of an unarticulated exoskeletal shield, and often is very simple in form. It is thought that at least some early protaspid larvae were likely to have been planktonic. The meraspid period is marked by a body with 2 or more articulated segments, and during the meraspid period, each molt meant the potential addition of usually one or two, but rarely greater numbers of articulated thoracic segments to the body of the growing trilobite. Several meraspid molts occurred, until the number of thoracic segments added to the meraspis achieved the number typical of the adult form of the species, and the general pattern of body morphology (shape and ornamentation) grew more similar to that of adults of the species. When the number of thoracic segments reached that of adult specimens, this marked the holaspid period, after which no further articulated segments were added with each molt. At this point, the growing animal is called a holaspis, and enters into the last period of development, in which the major change is not in form, but in increasing size. It is thought that most of the increase in size in the life cycle of a trilobite occurred during the holaspid period. As in some modern arthropods, there may have been a "terminal molt," after which the trilobite neither grew nor molted again. However, very large trilobites may have continually grown and molted, although less and less frequently over time.
Early period in which larva lacks articulated segments
Middle period in which juvenile bears articulated segments fewer in number than in the typical adult form
Adult number of articulated segments attained. Subsequent molts result in growth, but no additional thoracic segments.
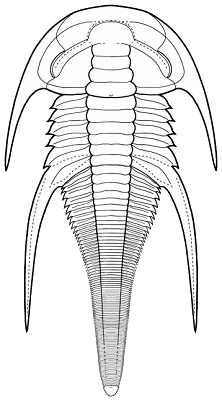
Addition of segments during development: When thoracic segments are added during the meraspid period, are they added to the growing juvenile from the cephalon or from the pygidium? In a trilobite with similar thoracic segments, it would be unclear whether new segments originate from the cephalon backward, or from the pygidium forward. The ontogeny sequence of the trilobite Shumardia (Conophrys) salopiensis (initially referred to as S. pusilla) provides a clear answer, because one of the thoracic segments bears long pleural spines. This reference segment is the fourth behind the cephalon in the holaspis, which has six thoracic segments (see adult holaspid specimen at left).
early holaspis 6 thoracic segments release of 2nd segment from pygidium forward
Trilobite Development Patterns
Full
articulations are bold vertical lines, unarticulated segments are unbolded and short, body tagmata (3) in colors. Key developmental milestones are marked by red arrows.
Patterns of development: Some modern arthropods, such as insects display remarkable differences in their development, moving through distinct phases (e.g., larva » pupa » adult) termed metamorphosis. Trilobite development was much simpler, though some phases are noted: typical development begins with an anamorphic mode (in which each molt adds articulated segments) but switches to an epimorphic mode (in which no more articulated segments are added per molt). This half anamorphic, half epimorphic mode is called hemianamorphic. The anamorphic mode is most easily seen in the meraspid period, while the holaspid development includes epimorphic growth. During the holaspid period, unarticulated segments might be added to the fused pygidium, but the total number of articulated units remains stable. This distinction between addition of body segments vs addition of articulations is important, because the timing of segment additions vs articulations is a major character that distinguishes developmental patterns in trilobites. Stabilization of developmental patterns: Evolution of trilobite body patterning during the long Paleozoic timescale reveals that the early Cambrian diversification involved more modification of the trunk region. Diversification in the Ordovician, by contrast, occurred within a narrower range of trunk conditions. By then, a premium on enrollment and protective strategies limited variation in the number of trunk segments. Trunk segment stability is seen in a number clades: e.g., the Asaphidae (8 thoracic segments), Trinucleidae (6 thoracic segments), Phacopoidea (11 thoracic segments), and Illaenina (10 thoracic segments).
The typical illaenine trilobite bear 10 holaspid thoracic segments.
The majority of trinucleioid trilobites bear 6 holaspid thoracic segments
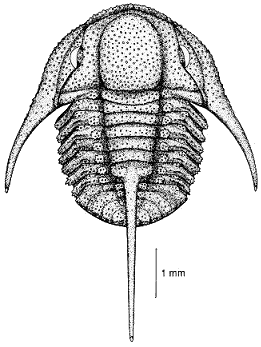
Mesonacis vermontanus, an olenelloid with a long post-macropleural prothorax of 12 segments, then a rather abrupt transition in segment size to the narrower opisthothorax.
Homonomy and Heteronomy: The majority of trilobites have homonomous (similar size/shape) thoracic segments. Some primitive trilobites in the Order Redlichiida (e.g., Olenelloidea and Emuelloidea) are notable for trunks bearing heteronomous (morphologically different) batches of segments (e.g., a small number of segments in a prothorax, typically ending with a macropleural segment, and followed by a large number of small segments making up an opisthothorax). There are exceptions, however (see Mesonacis vermontanus (left) with 16 segments in the prothorax (three of them pre-macropleural), and about 10 in the opistothorax).ns, however .. Among olenelloids and emuelloids, addition
of opisthothoracic segments may continue throughout the life of the trilobite, resulting in very large numbers of segments released from a tiny pygidium. The extreme development of this mode is seen in Balcoracania dailyi (in which 100+ segments may be achieved). With this many segments, it is thought that more than just one or two segments might be added during each molt. The opposite extreme is seen in the unusual trilobite Schmalenseeia fusilis, in which absolutely no articulations are created during the course of development, and the adult shows only a single, unarticulated dorsal shield, like a protaspis! .. The role of articulation between head and trunk in early arthropod evolution, and the idea that earliest arthropods may have been segmented but lack articulations, figure heavily in hypotheses about the ancestors of trilobites.
Balcoracania dailyi with >100 segments, the vast majority of these in a long opisthothorax.
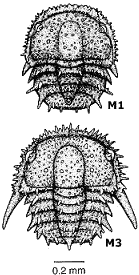
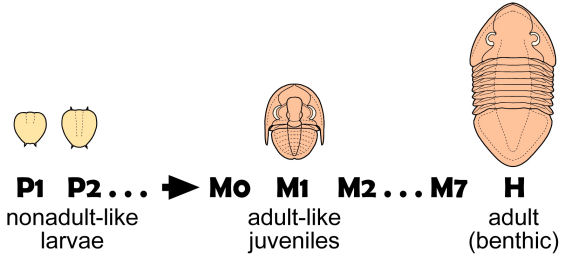
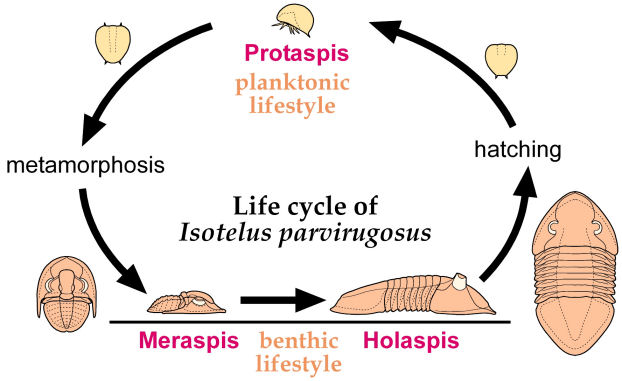
Ontogeny of Isotelus parvirugosus After Chatterton & Ludvigsen 1976 In the protaspid period (P1, P2, etc.) the larvae of Isotelus parvirugosus are not at all adult-like. However, an abrupt metamorphosis to adult-like form occurs in the meraspis (M0, M1, etc.) with growth in size and addition of thoracic segments.
Ontogeny, ecology and metamorphosis: Although in many trilobites growth is a relatively simple matter of increasing in size and adding segments until an adult holaspid form is achieved (as in the Shumardia example above), in several groups, such as the Asaphidae there may be two kinds of immatures: an "adult-like" juvenile form that is flat-bottomed and adapted to a benthic lifestyle, and a "non-adult-like" larval form that is globular, and thought to be adapted to a planktonic lifestyle. Typically, the nonadult- like larvae are seen earlier in ontogeny. Later (for example, during the meraspid period), the larval form undergoes metamorphosis into a juvenile form much more similar to the adult morphology.
How this change in form is thought to relate to changes in ecology during the life cycle is seen in the figure at left. Adults release eggs into the water column, and planktonic protaspid larvae bearing a globular shape emerge. In this case, all the protaspid stages are characterized by this effaced form. However, at the start of the meraspid period there is a metamorphosis, and juveniles settle out onto benthic habitats, and assume a benthic lifestyle and a much more adult-like form, until maturity is achieved and the cycle starts anew. In some trilobites, even meraspids may have a planktonic larval nonadult-like form. In others, adult-like protaspid juveniles are evident from early in development. Several different life history strategies have been defined, based on the presence of planktonic larvae and the timing of metamorphosis into adult-like form.
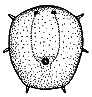
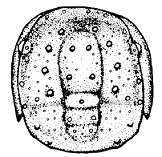
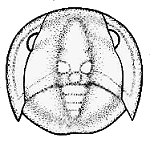
Protaspid stages: Most protaspid larvae begin simple, then assume more and more features in each molt before the meraspid period. Beecher (1895) named these sequential protaspid stages anaprotaspis (earliest stage), metaprotaspis (middle stage), and paraprotaspis (last, pre-meraspid stage). However, these stages were not initially well-defined, and only the first two are in current use. The anaprotaspid stage (see right) is typically simple, and lacks any clear transverse demarcation between protocephalon and trunk (protopygidium). There may or may not be any clear axis, and only cephalic lobes (four glabellar + one occipital) dominate the axial region, if any.
Anaprotaspides are usually simple globular or round shapes, though some marginal spines might be apparent. Even so, they sometimes bear distinguishing features that can indicate the order, suborder, or superfamily. For example, the asaphoid anaprotaspis (example top right) is globular, typically narrowing rearward, with up to three main pairs of submarginal spines, with a large, spiny hypostome, and no preglabellar field. Many brachymetopid anaprotaspides (example, right center) are circular, sometimes with an anterior spine and typically with two posterior spines with a incurved margin between them. Most known proetoid anaprotaspides (right bottom) are globular with three radiating pairs of short marginal spines.
Right: anaprotaspides of an asaphoid (Isotelus), brachymetopid (Dimeropyge), and proetoid (Cyrtosymbole).
asaphoid brachymetopid proetoid
Metaprotaspis stage: The metaprotaspis stage follows the anaprotaspis, and shows a transverse furrow (but not a true articulation) between the protocephalon and the trunk. Its axis consists of not only the glabella and occipital segment, but at least one segment posterior to those cephalic lobes. Thus these two distinguishing features: 1) a furrow marking the posterior margin of the cephalon, and 2) development behind the cephalon of segments or somatic region that will become the trunk; are together necessary to define the metaprotaspis.
In many groups the two events (transverse furrow and addition of postcephalic segments) occur together, but in some, there are post-cephalic segments apparent before any transverse furrow is developed. Therefore it is clear that the terms anaprotaspis and metaprotaspis cannot be clearly and consistently applied to the development of all trilobites. nor does the transition between the two stages occur at the same point in development. The terms can and are used for describing development of those groups where a clear distinction occurs, as in the Proetoidea (e.g., DeRosey-Aubril 2005), but should be used strictly descriptively and restricted to comparisons within related taxonomic groups with shared developmental patterns.
Right: metaprotaspides of a brachymetopid (Dimeropyge) and a proetoid (Proetus). Note the development of a transverse furrow between the protocephalon and the trunk, as well as post-cephalic trunk segments.
Dimeropyge Proetus
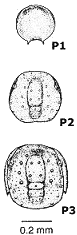
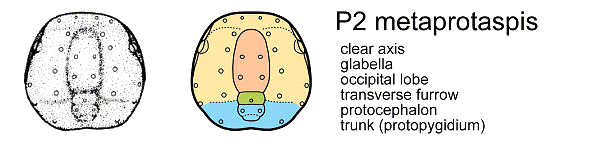
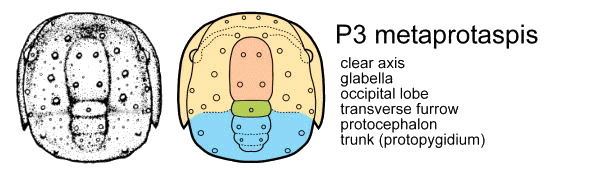
To summarize and clarify the key features of the anaprotaspis and metaprotaspis, these diagrams show the salient differences, color coded, for the protaspid stages of Dimeropyge speyeri.
The P1 stage (top) shows facial sutures, but no sign of an axis and no transverse furrow separating regions of the larva. It clearly qualifies as an anaprotaspis.
The P2 stage (middle) shows a clear axis, which is further color coded in orange (glabella) and green (occiptial lobe) for the cephalic lobes, and blue for the trunk axial segments. The first sign of a transverse furrow appears, extending from the occipital lobe, separating protocephalon (yellow) from trunk (blue).
The P3 metaprotaspis stage (bottom) adds more segments to the trunk, and the librigenae (free cheeks) now show short genal spines. Where the base of the genal spine meets the facial suture is the junction between protocephalon and trunk.
The concept discussed earlier, of two general forms of protaspis -- an "adult-like" form with recognizable cephalic and post-cephalic portions, and a "nonadult-like" form that is simple, smooth, and globular -- seems related to the anaprotaspis and metaprotaspis in most groups. There is often a metamorphosis between the earliest nonadult-like protaspid instar(s) and the later adult-like protaspides, leading Chatterton & Speyer (1997) to suggest that the metaprotaspis may be a synonym for the adult-like protaspis. Indeed, where the transition from nonadult-like larva to adult-like juvenile occurs between protaspis and meraspis periods, there is technically NO metaprotaspid stage. Such is the case with many Asaphida.
A Few Protaspides of Different Trilobite Orders This sampler merely hints at the diversity of protaspid forms
Agnostida
Redlichiida
Corynexochida
Lichida
Odontopleurida
Phacopida
Proetida
Asaphida
Ptychopariida
Ontogeny and classification: Ontogeny plays an important role in assessing the higher classification of trilobites (e.g., Fortey 1990). There is a great deal of variation of form among the protaspid and meraspid larvae of the different trilobite orders, and related families and suborders show similar immature forms. For example, most of the Asaphida show a consistent protaspid form (called an asaphoid protaspis), and this ontogenetic similarity links groups that look quite different in their mature forms (for example, the Asaphida superfamilies Trinucleioidea and the Asaphoidea). Similarly, although the three suborders of Phacopida (Calymenina, Cheirurina, and Phacopina) are quite different in their holaspid forms, they share a number of similarities during their development that are considered synapomorphies (shared derived characters) indicating monophyly (descent from a common ancester). Similar arguments were made (Fortey 1997) to unite the Lichidae and Odontopleuridae within the Lichida, though in that case, there are some ambiguities and controversy. Fortey & Owens' (1975) distinction of the Proetida as an order separate from the Ptychopariida was also partly based on the protaspid establishment of adult-like cephalic characteristics, such as a tapering glabella and a preglabellar field (as in the proetid protaspid specimen depicted above). Similarly, Lee & Chatterton (2003) showed that among the protaspides of Corynexochida, hexagonal form (above) unites members of suborders Leiostegiina, Illaenina, and Corynexochina. Immature stages of trilobites are not often preserved with sufficient detail to assist such systematic analyses. This hinders progress in clarifying long-standing questions of relationships between trilobite orders (Fortey 2001).
Recent work on the ontogeny of Proetida by Rudy Lerosey-Aubril and others has resulted in advances in our understanding of the superfamilies of the Proetida, and comprehensive compilations of the literature on these and other trilobite ontogenies. I heartily encourage interested readers to consult his webpages on The Ontogeny of Trilobites.
Selected Bibliography on Trilobite Ontogeny
Beecher, C.E. 1895. The larval stages of trilobites. The American Geologist. 16:166-97.
Chatterton, B.D.E., and S.E. Speyer. 1990. Applications of the study of trilobite ontogeny. Short Courses in Paleontology, Paleontological Society 3:116-36.
Chatterton, B.D.E., and S.E. Speyer. Ontogeny, pp 173-247 in Kaesler, R. L., ed. 1997. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. xxiv + 530 pp., 309 figs. The Geological Society of America, Inc. & The University of Kansas. Boulder, Colorado & Lawrence, Kansas.
Fortey, R.A. 1990. Ontogeny, hypostome attachment, and trilobite classification. Palaeontology 33:529-76,
figs. 1-19, pl. 1.
Fortey, R. A. 2001. Trilobite systematics: the last 75 years. J. Paleontology. 75(6):1141-51.
Fortey, R.A. & N.C. Hughes. 1998. Brood pouches in trilobites. J. Paleontology 72(4):638-49.
Fortey, R.A. & R.M. Owens. 1975. Proetida: a new order of trilobites. Fossils and Strata 4:227-39.
Hughes, N.C. 2003. Trilobite tagmosis and body patterning from morphological and developmental perspectives. Integrative and Comparative Biology 43(1):185-206
Hughes, N.C. 2007. The evolution of trilobite body patterning. Annual Review of Earth and Planetary Sciences. 35:401-34.
Hughes, N.C., A. Minelli & J. Fusco. 2006. The ontogeny of trilobite segmentation: a comparative approach. Paleobiology 32(4):602-27.
Lee, Dong-Chan & B.D.E. Chatterton. 2003. Protaspides of Leiostegium and their implications for membership of the order Corynexochida. Palaeontology46(3):431-46.
Lerosey-Aubril, R. and R. Feist. 2005. First Carboniferous protaspid larvae (Trilobita). J. Paleontology, 79(4): 702-718.
Stubblefield, C. J. 1926. Notes on the development of a trilobite, Shumardia pusilla (Sars). Zool. J. Linnean Soc, 35:345-372.
Whittington, H.B. Ontogeny of Trilobita, pp O127-O145 in Moore, R.C., ed. 1959. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. Geological Society of America & University of Kansas Press. Lawrence, Kansas & Boulder, Colorado. xix + 560 pp., 415 figs.
Zhang, X. & B. Pratt. 1994. Middle Cambrian arthropod embryos with blastomeres. Science 266:637-9.