Trilobite Enrollment
last revised 13 December 2008 by S.M. Gon III
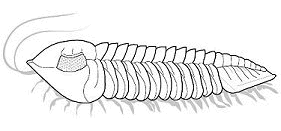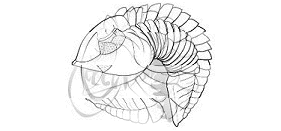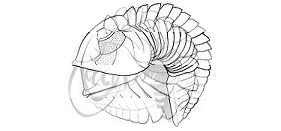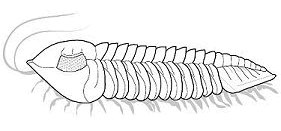
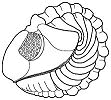
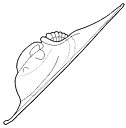
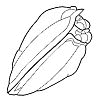
What is enrollment? Most trilobites could enroll
into a defensive ball or capsule, via the flexible articulation of the thoracic
segments, bringing the cephalon and pygidium together in a
protective closed capsule that shielded the antennae, limbs, and soft ventral
surface. While in that enrolled state, the trilobite could watch and wait
until conditions were safer (as in the example specimen of Phacops to the left). Some groups of trilobites (e.g. the Phacopina) developed specialized morphological features that aided enrollment, called coaptative structures. These were complementary morphological features that allowed close interlocking of opposing surfaces (coaptation). The cephalon and pygidium
of enrolled trilobites often have similar shapes that allow a tight match,
even to the point of special notches that fit the edges of enrolled thoracic
segments and the pygidial border (see below).
In general, trilobites enrolled by contracting internal muscles,
bending the flexible integument (shell) between each of the rigid
thoracic segments so that the cephalon and pygidium were brought
together, and the thoracic pleurae slid into an overlapping radial
pattern. Some modern arthropods, such as crustacean isopods, are
similarly able to enroll into very compact, sphereical capsules that
are resistant to their typical enemies. The animated example above
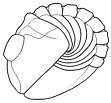
shows the progressive enrollment of a phacopid trilobite: Acaste downingiae. Notice how the limbs
and antennae are tucked in to fit within the enrolled exoskeleton, and how
in the fully enrolled state, the pygidium and several thoracic pleurae contact
the cephalon and no limbs or ventral surfaces are exposed at all. Where
the pygidium and thoracic pleurae make contact with the cephalon, there
is sometimes a specialized coaptative structure, called the vincular furrow (from the latin vinculus, a locking or binding device), which matches the shape of the
pygidial margin and the ends of the thoracic pleurae (see below).
...
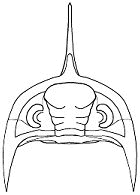
frontal lobe >
of glabella
vincular >
furrow
vincular >
notches
Vincular furrow and notches
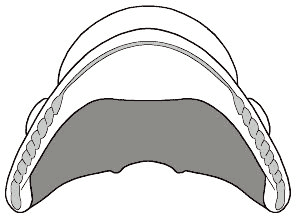
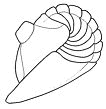
Here is the underside of the cephalon of the phacopine trilobite Paciphacops.
When enrolled, it would appear rather similar to the enrolled Phacops
at the top of this page. The dark grey area is the large concavity
of the cephalon, where anterior organs and limbs would be. The lighter grey
narrow arch-shaped feature is the vincular furrow, and the vincular
notches are the rope-like scalloped lobes arranged at the bottom left and right
of the vincular furrow. The furrow accomodates the edge of the pygidium on
enrollment, and the notches accomodate the rounded ends of the thoracic pleural
segments, forming a near-perfect, tight fit (coaptation). One rounded thoracic
pleural termination would fit into each of the vincular notches (see the animated
enrollment of Acaste above)
More enrollment examples
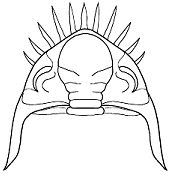
Genal and pygidial spines are often designed to offer extra protection
when the trilobite is in an enrolled state. Below are the top views
of two extended and enrolled phacopine trilobites. Note how the cephalic and pygidial
spines offer defense when the animal is in the enrolled state.
When Dalmanites (above) enrolled (right), an unwieldy triangle
of spines was created to present to would-be predators.
When Comura (above) enrolled (right), the result was a very
discouraging fan of spines.
Types of enrollment: Both the 1959
and 1997 Treatises devoted discussions to enrollment and
coaptative structures of trilobites, noting that by virtue of
articulation of the thoracic segments, most if not all trilobites could
curl the exoskeleton to maximize protection of the vulnerable ventral
surfaces and limbs. It is interesting that the most primitive
trilobites (e.g., most Olenelloidea) did not have well-developed
coaptative structures and could not form closed enrolled capsules. Once
enrollment was well established during the Cambrian, post-Cambrian
evolution of trilobite exoskeletons seemed restricted by the need to
maintain effective enrollment. The 1959 Treatise and Bergstrom's 1973
review of
enrollment described some general types of enrollment. These go as far back as Barrande (1852) in which three main types of complete enrollment were named: sphaeroidal, double, and discoidal.
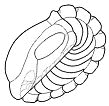
Sphaeroidal enrollment
occurs when all of the thoracic segments participate more of less
equally in curling the body of the trilobite onto itself. This was
described as the most common type of enrollment, especially among
isopygous and macropygous trilobites. Agnostida, isopygous trilobites
with only two or three segments, nonetheless bring the cephalon and
pygidium tightly together, and so qualify as sphaeroidal enrollers.
Similarly, enrolling trilobites with relative few segments, such as the
Asaphoidea (ca 8 segments) and Phacopoidea (ca 10 segments) generally
involve all segments more or less equally in enrollment, and are
considered sphaeroidal enrollers (see example Nyterops,
right). In fact, the stabilization of segment number in these clades
may be due to the strong adaptive advantage of effective enrollment.
Variations among sphaeroidal
enrolling trilobites include ones in which either the cephalon or the
pygidium extend further than the other when enrollment is complete
(e.g., Pseudomegalaspis, below).
Double enrollment
occurs when the pygidium and hindmost thoracic segments are hidden
under the front edge of the cephalon. Seen in primitive Cambrian
micropygous trilobites such as Ellipsocephalus, Bergstrom (1973) referred to this kind of enrollment as spiralenrollment.
Discoidal enrollment
involves flexing of only the anterior portion of the thorax, while the
rest of the thorax and pygidium behaves like a closing lid. This type
of enrollment is seen in micropygous trilobites with cephalic
development, such as harpetids (example left) and trinucleioids, and was referred to
as basket & lidenrollment by Bergstrom (1973).
Incomplete enrollment
occurs when the thoracic pleurae do not completely enclose the ventral
surfaces at full enrollment, leaving a ring of pleural spines
projecting around a lateral gap. This is the case not only with
primtive spinose trilobites such as olenelloids, and paradoxodoids, but
is seen in more advanced trilobites such as Selenopeltis and some cheirurids. Bergstrom referred to this as cylindrical enrollment.
Some trilobite workers, such as Bergstrom (1973), suggested that the
different kinds
of enrollment might be useful in classification of trilobites. However,
there
is a great deal of variability in trilobite enrollment, and other
workers suggest that there is so
much inconsistency that the utility of enrollment patterns among
trilobites for higher level classification is relatively low, or so
tied to other obvious morphological features as to be redundant. By the
1997 Treatise, the general view was that the basic enrollment types
described were of limited use, and that enrollment was "not a character
to be considered fundamental in classification."
Although of uncertain value for trilobite classification, enrollment
remains a fascinating feature of trilobite morphology and worth
understanding. Here are a few more images of the enrolled form of
various species of trilobites. The different types of enrollment have
been labeled, although they are not consistently used.
Notice how in the case of Ellipsocephalus and Harpes, there
is not an exact fit between the cephalon and the enrolled thorax and pygidium.
Faint lines show how the thorax and pygidium is tucked within the concavity
of the cephalon in those cases. In Pseudomegalaspis, there is an
"overbite" that suggests that a portion of the pygidial venter is exposed,
but in this case, a very wide pygidial doublure serves as a shield, and
there are actually no exposed limbs or soft ventral surface. Finally, below
are two images of enrolled trilobites, demonstrating the elegant beauty
of the protective capsule formed by enrollment.
A beautiful example of enrollment in the trilobite Pliomera, family Cheiruridae, Order Phacopida
Enrolled Illaenus bayfieldi,
family Illaenidae, Order Corynexochida
Selected references:
Barrande, J. 1852. System Silurien du Centre de la Boheme. I.
Recherches Paleontologiques, vol. 1 (Crustaces: Trilobites) Prague
& Paris. xxx + 935 pp., 51 pl.
Bergstrom, J. 1973. Organization, life, and systematics of
trilobites. Fossils and Strata 2:1-69.
Clarkson, E.N.K., & H.B. Whittington. 1997. Enrollment and coaptative structures. pp. 67-74 in: R.L. Kaesler, ed. Treatise on Invertebrate Paleontology. Part O: Arthropoda 1, Trilobita, Revised. Lawrence, Kansas.
Harrington, H.J. 1959. Enrollment. pp. O102-O107 in: R.C. Moore, ed. Treatise on Invertebrate Paleontology. Part O: Arthropoda 1. Lawrence, Kansas.
 What is enrollment?
What is enrollment?