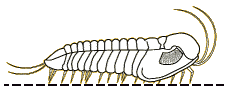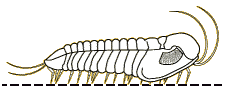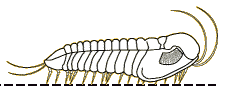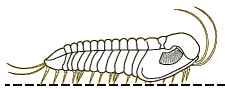
click image for
pictorial guide Kettneraspis
 | ORDER ODONTOPLEURIDA Whittington 1959
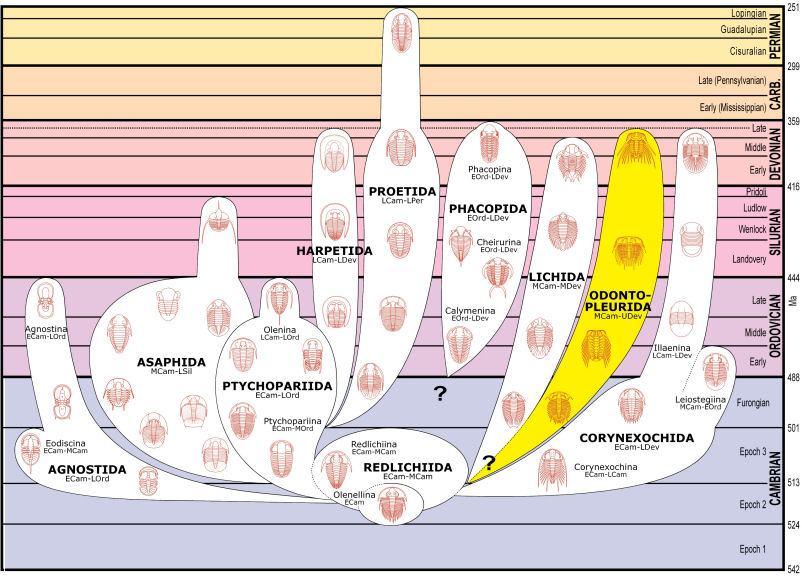
last revised 09 JULU 2020 by S. M. Gon III Introduction: distinctive group of typically very spinose trilobites with densely granulate or tuberculate exoskeletons; alternately placed as superfamily Odontopleuroidea in a broad concept of Order Lichida.
Cephalon: opisthoparian sutures; glabella extending to anterior border, lobation complex in advanced forms; simple holochroal eyes, usually not large; conterminant hypostome, not particularly large.
Thorax: variable, 8-13 segments, usually spine-tipped, sometimes greatly developed with distinctive spine patterns (e.g., Odontopleuroidea).
Pygidium: micropygous to subisopygous, with multiple pairs of spines, even in primitive forms (Dameselloidea).
Other: doublure not extensive, lacking terrace ridges
Occurrence: Middle Cambrian to Devonian (Frasnian)
Superfamilies: Odontopleuroidea, Dameselloidea. |
Dicranurus
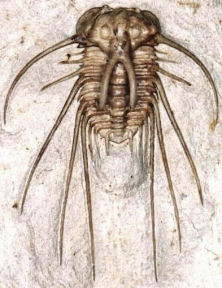
 | Superfamily Odontopleuroidea Introduction: typically very spinose and densely sculptured trilobites.
Cephalon: convex; glabella tapering forward or subparallel, extending to anterior margin or nearly so, less complex lobation than in sister order Lichida; eye ridges run from anterior end of glabella to palpebral lobe; opisthoparian sutures, often placed on sutural ridges; distinct notch in margin of free cheek adjacent to where anterior sutures intersect cephalic margin; facial sutures secondarily lost in some genera; genal spines typically present.
Thorax: 8 – 10 segments; tips of each bear 2 – 3 pairs of spines (anterior pair often difficult to see, ventrally directed); often with symmetrical rows of pleural spines or tubercles.
Pygidium: micropygous, short, transverse, with 2 – 3 axial rings (3rd often faint), one or more pairs of tubular border spines, the largest of which connected to first axial ring by prominent ridge.
Occurrence: Upper Cambrian to Upper Devonian (Frasnian)
Families: Odontopleuridae
Genera: Odontopleuridae: Acanthalomina, Acidaspidella?, Acidaspides?, Acidaspis, Anacaenaspis (=Bruxaspis), Apianurus, Archaeopleura, Boedaspis, Borkopleura, Brutonaspis, Calipernurus, Ceratocara, Ceratocephala (=Bounyoungia; =Onchaspis; /Trapelocera), Ceratocephalinus, Ceratonurus, Chlustinia, Dalaspis, Diacanthaspis, Dicranurus, Dudleyaspis, Edgecombeaspis, Eoleonaspis (=Bojokoralaspis), Exallaspis, Gaotania, Globulaspis, Hispaniaspis, Isoprusia (=Mauraspis), Ivanopleura, Kettneraspis (=Grossia), Koneprusia, Laethoprusia, Leonaspis (=Acanthaloma), Meadowtownella, Miraspis (=Elbaspis), Ningnanaspis, Odontopleura, Orphanaspis, Periallaspis, Primaspis, Proceratocephala (=Drummuckaspis), Radiaspis (=Xanionurus; =Charybdaspis), Rinconaspis, Selenopeltis (=Languedopeltis; =Polyeres), Selenopeltoides, Sinespinaspis, Stelckaspis, Taemasaspis (=Gondwanaspis; =Snoderaspis), Uriarra, Whittingtonia. |
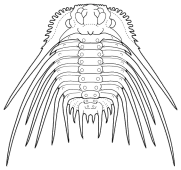
Damesella
 | Superfamily
Dameselloidea Introduction: primitive Odontopleurida, surface finely to coarsely granulose.
Cephalon: opisthoparian sutures; glabella tapering forward, less complex lobation than in Odontopleuroidea; genal spines typically present.
Thorax: with up to 13 segments, less specialized than in Odontopleuroidea, more as in typical polymerids, but sometimes with extended spines.
Pygidium: longer pygidia than in Odontopleuroidea, with more axial segments; tapering axis, thicker-set marginal spines, 1-7 pairs of pleural pygidial spines of varying length
Occurrence: Middle Cambrian to Upper Cambrian(?)
Family: Damesellidae
Genera: Damesellidae: ?Adelogonus, Ariaspis, Bergeronites (=Spinopanura), Blackwelderia (=Parablackwelderia), Blackwelderioides, Chiawangella, Cyrtoprora, Damesella (=Hybowania; =Eodamesella), Damesops (=Meringaspis; =Paradamesops), Dipentaspis, Dipyrgotes, Drepanura, Duamsannella, Fengduia, Guancenshania, ?Hercantyx, Histiomona, Jiawangaspis, Liuheaspis, Metashantungia, Neodamesella, Palaeadotes, (=Pseudobergeronites), Paradamesella (=Falkopingia), Parashantungia, Pingquania (=Oxygonaspis), Pionaspis, Protaitzehoia, Pseudoblackwelderia, Shantungia, Stephanocare, Taihangshania, Taitzehoia, Teinistion (=Dorypygella), Xintaia, Yanshanopyge. |
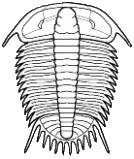
Selenopeltis
Miraspis
 | ADDITIONAL CLASSIFICATION NOTES FOR ODONTOPLEURIDA:
This treatments recognizes an Order Odontopleurida separate from Order Lichida, although protaspid similarities have been used to suggest the two may ultimately be related (Thomas & Holloway 1988); both lichid and odontopleurid protaspides possess distinctive paired spine or tubercle patterns on cephalon and protopygidium, have an anterior border, distinct axis, and marginal spines on protopygidium (Thomas & Holloway 1988). Both Lichidae and Odontopleuridae are spiny, bear tubercles as prosopon, and are marked by complex glabellar lobation. Largely on these bases, the family Odontopleuridae was afforded by Fortey (1997) superfamily status (Odontopleuroidea) and included in the Order Lichida.
However, several lines of evidence argue for separate ordinal status for odontopleurids. Whittington (1956) points out that the lichid protaspis is twice the size of those of odontopleurids. Odontopleurid protaspide hypostomes also lack slits on the lateral hypostomal border borne by those of lichids (and styginids). An analysis of lichid protaspides by Campbell and Chatterton (2009) place Odontopleurida farther from lichids than styginids and proetids, supporting an argument that Odontopleurida are not the sister group to Lichida. Despite both groups being ornamented with tubercles and typically spiny, the lobation of the glabella, complex in both, show some major differences in origin (lichid lobes arising from the axial furrow, while odontopleurid lobes arise via standard glabella development (Tripp & Evitt 1981). Whittington (2002) asserts that the holaspids of Odontopleuridae and Lichidae also differ in several other respects, including ornamentation (primarily granules and tubercles in lichids, spines in odontopleurids), extent of ventral calcification (extended doublure with terrace ridges in lichids, not developed in odontopleurids), and hypostome size (larger in lichids).
The primitive family Lichakephalidae as recognized by Jell & Adrain (2003) includes genera that have affinities to Lichidae and others to Odontopleuridae, and is pointed to as further evidence for relationship between the two families. However, some workers (e.g., Shergold et al 2000) do not accept the synonymy of Eoacidaspididae and Lichakephalidae. Examination of Cambrian lichakephalid/eoacidaspidid genera Acidaspides and Acidaspidella (Bruton 1983) suggest they should be assigned to Odontopleuridae. If the Lichakephalidae is polyphyletic, then not all of its members should be included in Lichoidea, but placed in Odontopleuridae (marked with "?" above).
Fortey (1990) included the M-U Cambrian Damselloidea as a sister group to Odontopleuroidea, citing several uniting characters: 1) narrow, ledglike anterior cranidial border, against which glabella abuts sharply, 2) deeply scribed, inflated eye ridges, 3) transverse hypostome with wide posterior border, 4) spinosity, especially on pygidium and anterolateral margins of free cheeks, 5) occipital tubercle without thoracic homologs 6) 3rd glabellar lobe reduced or absent. Presence of Acidaspides praecurrens and 'odontopleurid protaspides' in Upper Cambrian Kazakhstan attributable to dameselloids further strengthen this relationship (Fortey pers com 2007).
Several post-2000 publications have recognized the order Odontopleurida in systematics listings (e.g, Calner et al 2006, Chatterton & Ludvigsen 2004, Chatterton et al 2006) further validating, via usage, the ordinal status of Odontopleurida.
Click here, for order Lichida split from the odontopleurid-damesellid clade.
Click here for the concept of an order Lichida that includes odontopleurids, damesellids, and lichids.
Expanded and complete genera listings for the families above modified from Jell & Adrain (2003).
LITERATURE CITED:
Bruton, D. 1983. Cambrian origins of the odontopleurid trilobites. Palaeontology 26(4):875-85.
Calner, M., P. Ahlberg, N. Axheimer &L. Gustavsson. 2006. The first record of Odontopleura ovata (Trilobita) from Scandinavia: part of a middle Silurian intercontinental shelly benthos mass occurrence. GFF 128: 33-37. Campbell, M.J. & B.D.E. Chatterton. 2009. Silurian lichid trilobites from northwestern Canada: Ontogeny and phylogeny of lichids. J. Paleontol. 83(2):263-79.
Chatterton, B., R. Fortey, K. Brett, S. Gibb & R. McKellar. 2006. Trilobites from the upper Lower to Middle Devonian Timrhanrhart Formation, Jbel Gara el Zguilma, southern Morocco. Paleontographica Canadiana 25:1-177., 54 pls.
Chatterton, B.D.E. & R. Ludvigsen. 2004. Early Silurian trilobites of Anticosti Island, Quebec, Canada. Palaeontologica Canadiana 22:1-264. Fortey, R.A. 1990. Ontogeny, hypostome attachment, and trilobite classification. Palaeontology 33:529-76.
Fortey, R.A. 1997. Classification. In Kaesler, R. L., ed. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. xxiv + 530 pp., 309 figs. The Geological Society of America, Inc. & The University of Kansas. Boulder, Colorado & Lawrence, Kansas.
Jell, P.A. & J.M. Adrain. 2003. Available generic names for trilobites. Memoirs of the Queensland Museum 48(2):331-553.
Shergold, J.H., R. Feist & D. Vizcaino. 2000. Early Late Cambrian trilobites of Australo-Sinian aspect from the Montagne Noir, Southern France. Palaeontology 43(4):599-632.
Thomas A.T. and D.J. Holloway. 1988. Classification and phylogeny of the trilobite order Lichida. Phil. Trans. Royal Soc. London 321:179-262.
Whittington, H.B. 1956. Beecher's lichid protaspis and Acanthopyge consanguinea (Trilobita). J. Paleontol. 30:1200-04.
Whittington, H.B. 2002. Lichidae (Trilobita): Morphology and classification. J. Paleontology 76(2):306-20. |