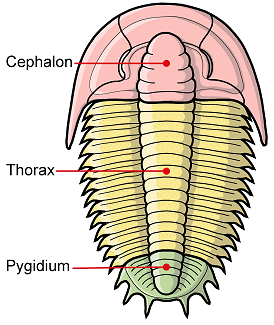
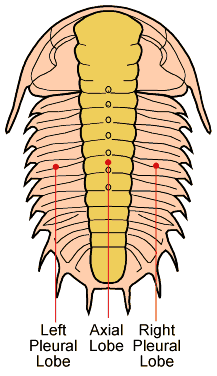
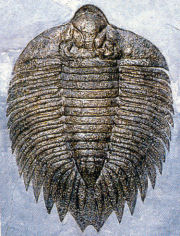
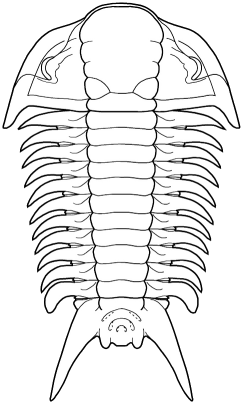
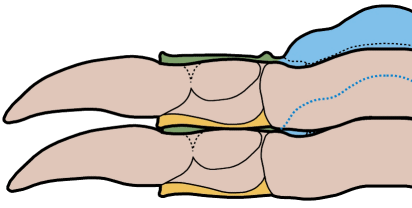
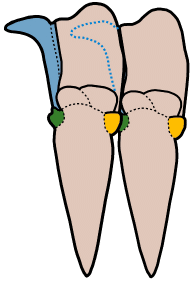
In the images above, sometimes the similarity between the occipital segment and the thoracic segments are striking. In Reedops deckeri, round lateral lobes on either side of the thoracic axis are mirrored in not only the occipital lobe, but also the glabellar lobe immediately anterior to it. The occipital segment of Modocia laevinucha is partially defined by the posterior facial suture, which further enhances the similarity between that cephalic segment and the first thoracic segment. In Huntoniatonia
oklahomensis, the pygidial segments look very much like fused thoracic segments. The spines of the pygidium of Arctinurus
boltoni are so similar to the backward-facing thoracic pleural spines that it is difficult to tell where the thorax ends and the pygidium begins (there are 11 thoracic segments and three pairs of pygidial spines).
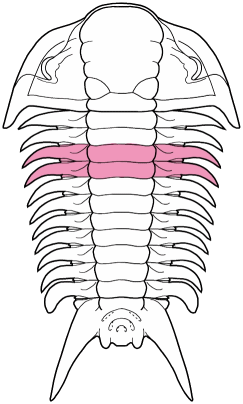
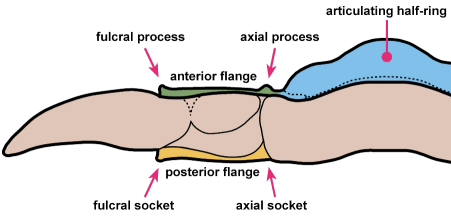
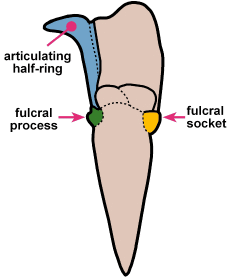
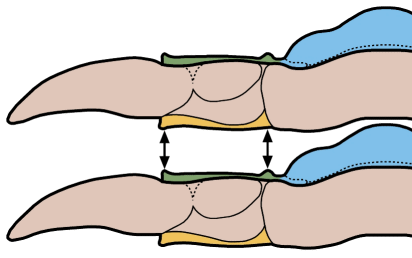
A structural thoracic feature associated with enrollment is the pleural fulcrum, where there is a clear change of angle from the near-horizontal inner pleural region to a more steeply-sloped outer region. In the trilobites above, the fulcrum is particularly apparent in Calymene, Reedops, and Breviscutellum, and is subtle or lacking in Olenellus, a primitive species that was not thought to enroll well.
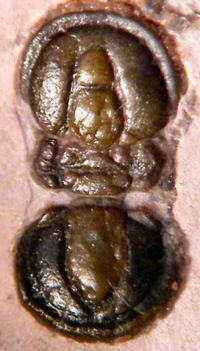
The minimum number of articulated thoracic segments in a typical trilobite with articulations between cephalon, thorax and pygidium is two, in
Agnostida such as
Peronopsis interstricta (below). However, one strange trilobite,
Schmalenseeia
fusilis, has no articulations at all! Cephalon, thorax, and pygidium are all fused into an unbending oval dome, and it is only the presence of articulated close relatives that this strange species can be called a trilobite at all! Typical trilobites might have several to many thoracic segments (
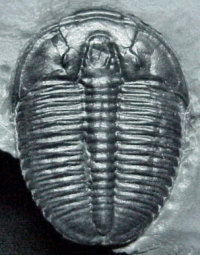
Elrathia kingi, perhaps the most common trilobite known, has 13 articulated thoracic segments). The maximum number of articulated thoracic segments is seen in the
emuellid trilobite
Balcoracania dailyi, with an exceptional 100+ articulated thoracic segments!
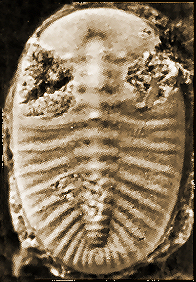 |  |  |  |
Schmalenseeia
fusilis
0 segments | Peronopsis
interstricta
2 segments | Elrathia
kingi
13 segments | Balcoracania
dailyi
100+ segments |
Lateral
articulation
Although
the primary bending plane of a trilobite was vertical, some horizontal thoracic bending was also possible. This added flexibility might be useful for foraging through complex environments, such as reefs. However, the majority of trilobite specimens are more or less straight, with little or no lateral axial flexing, although many are found showing vertical flexing, including full enrollment. Perhaps lateral bending was limited, or in death relaxation of internal muscles straightens the body. A few long-bodied primitive species have been found showing lateral flexing, as in the Balcoracania dailyi (left), Olenellus fowleri (middle), and Olenellus terminatus (right) shown here. wide pleural fields, relative to the axis, would have greatly limited lateral range of motion in a trilobite such as Elrathia (above).
Thanks to George Ast for the Nevadia and Olenellus images.
| 
Balcoracania daily
| 
Olenellus fowleri
| 
Olenellus terminatus
|
Some
references:
Chatterton,
B.D.E. & M. Campbell. 1993. Enrolling in trilobites: A review and some new characters. Memoirs of the Association of Australian Paleontologists 15:103-23.
Harrington, H.J. 1959. General description of Trilobita: Thoracic region. In: Moore, R.C., ed. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. Geological Society of America & University of Kansas Press. Lawrence, Kansas & Boulder, Colorado. xix + 560 pp., 415 figs.
Hughes, N.C. 2003. Trilobite tagmosis and body patterning from morphological and developmental perspectives. Integrative and Comparative Biology 43(1):185-206.
Hughes, N.C. 2007. The evolution of trilobite body patterning. Annual Review of Earth and Planetary Sciences. 35:401-34.
Whittington, H.B. 1997. Morphology of the exoskeleton. In: Kaesler, R. L., ed. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. The Geological Society of America, Inc. & The University of Kansas. Boulder, Colorado & Lawrence, Kansas. xxiv + 530 pp., 309 figs.
all figures above ©1999-2007 by S.M.
Gon III created with Macromedia Freehand

Walking Trilobite animation ©2000 by S. M. Gon III